Ornithomimus ("imitador de aves") es un género conocido por dos especies de dinosaurios terópodos ornitomímidos, que vivieron a finales del período Cretácico, hace aproximadamente entre 70 y 65 millones de años, en el Mastrichtiano, en lo que hoy es Norteamérica. Ornithomimus velox, la especie tipo, fue nombrada en base a un pie y una mano proveniente de la Formación Denver, pero mejor material ha sido encontrado en Canadá, incluyendo a Ornithomimus edmontonicus un excelente especimen articulado del Parque Provincial del Dinosaurio, en la Formación Cañón Herradura. Otros especimenes asignados a Ornithomimus han sido encontrados en la costa este de los Estados Unidos. El primer fósil de Ornithomimus fue encontrado en 1889 y nombrado por el paleontólogo norteaméricano Othniel C. Marsh en 1890.Todas las especies de ornitomímidos poseían un pequeño cráneo y huesos ligeros, lo cual sugiere que eran ágiles y alcanzaban altas velocidades. Su pico sin dientes les permitía comer diversos tipos de alimentos, con lo que probablemente eran omnívoro, alimentándose de hojas, frutos, mamíferos y reptiles pequeños, mariscos, insectos, etc. Llegó a medir 3,5 metros de largo, 2 de alto y a pesar 150 a 200 kilogramos. Tenía un cuello largo, un cráneo similar a de una ave que terminaba en un pico, pies con tres dedos y `largos y delgados brazos. Difiere de otros ornitomímidos, como Struthiomimus, por tener unas delgadas y largas garras en manos y pies y en tener metacarpales y dedos de longitudes similares.[1] Estas manos son muy similares a los de un perezoso, lo que llevo a sugerir a Henry Fairfield Osborn que eran usadas como ganchos para la alimentación.
[editar] En la cultura popular
Ornithomimus ha tenido una prominente ubicación en la serie para televisión Prehistoric Park, donde aparecían, en mayor o menor medida, en cada episodio. Fueron retratados como animales de manadas cuyas crías tenían plumas e impronta con la primera cosa que veían. También se los veían teniendo hábitos de alimentación más similares al de un pato que a una avestruz, un comportamiento basado en una hipótesis hoy refutada[2] en la cual se los consideraba como animales filtradores similares a los actuales flamencos.[3] Ornithomimus además apareció en un gran número de películas, incluyendo Fantasía (1940), El valle de Gwangi (1969), Planet of Dinosaurs (1978) y en el film de IMAX T-Rex: Back to the Cretaceous (1998).
Amargasaurus ("reptil de Amarga") es un género representado por una única especie de dinosaurio saurópodo dicreosáurido, que vivió a principios del período geológico Cretácico, hace aprimadamente 130 millones de años, en el Barremiano, en lo que hoy es Sudamérica. Amargasaurus se encuentra cercanamente emparentado con el más antiguo dinosaurio afrícano Dicraeosaurus. Al igual que este presenta grandes espinas neurales bífidas, aunque en Amargasaurus son de mayor tamaño.El amargasaurio medía 10 m de longitud, 4 m de altura y pesaba 8 t, siendo pequeño para un saurópodo. Habría sido un herbívoro cuadrúpedo con un cráneo largo y bajo en el extremo de un cuello relativamente corto, como en su cercano pariente Dicraeosaurus. Poseía unas altas espinas, más alta que en otros saurópodos, que se bifurcaban y nacían desde las vértebras. Estas espinas se extendían desde la nuca, por el cuello hasta la mitad de la espalda. Es posible que sostuvieran una vela y suele ser representado con esta aunque esto es rechazado por Gregorio S. Paul en 2000.[1] Otra posibilidad es que estuvieran recubierta de grasa o tejido muscular.
Esqueleto de Amargasaurus montado en el Museo de Melbourne.La característica más obvia del esqueleto de Amargasaurus es la serie de espinas dorsales altas en el cuello y las vértebras de la espalda. Las espinas dorsales son las más altas en el cuello, donde se aparean en dos filas paralelas. Estas filas continúan a lo largo del lomo, disminuyendo en altura mientras que se acercan a las caderas. Las últimas vértebras dorsales y sacras muestran solamente una espina, que son largas pero mucho más cortas que las del cuello, comparables a otros saurópodos. Estas espinas dorsales pudieron haber apoyado un par de velas altas de la piel. Las velas similares se ven en los dinosaurios sin relación Spinosaurus y Ouranosaurus, así como los pelicosaurios Dimetrodon y Edaphosaurus. Hay una variedad de hipótesis para la función de estas velas, incluyendo defensa, la comunicación (para los propósitos de apareamiento o para el simple reconocimiento de la especie) o la regulación de temperatura. Sin embargo, su función verdadera sigue siendo desconocida. Las espinas dorsales similares se encuentran en las vértebras presacras de Dicraeosaurus de África, aunque no casi como altas.
[editar] Historia
Amargasaurus, con la "vela de piel".El nombre Amargasaurus fue colocado en 1991 por los paleontólogos argentinos Leonardo Salgado y José Bonaparte,[2] debido de que sus restos fueron encontrados a la orilla del Arroyo La Amarga en Neuquén provincia de Argentina. La Amarga es también el nombre de una localidad cercana y de la formación geológica donde fue encontrado. El nombre especifico, ' ' A. cazaui' ' se le coloco en honor del hombre que descubrió el sitio, el Dr. Luis B. Cazau, geólogo de YPF, que en ese entonces propiedad del gobierno. Este sitio está situado en las secciones más bajas de la Formación La Amarga, que data en el Barremiano al Aptiano temprano, durante el Cretácico inferior, hace alrededor de 130 a 120 millones de años. Una réplica del esqueleto de este dinosaurio se encuentra en el Museo Argentino de Ciencias Naturales Bernardino Rivadavia.
Amargasaurus es conocido por un relativamente completo único esqueleto. Este esqueleto incluye la base del cráneo, todas las vértebras del cuello, espalda y caderas, así como una porción de la cola. El hombro derecho también se conoce, al igual que el miembro superior e inferior izquierdo, y el ilion izquierdo, un hueso de la pelvis.
[editar] Clasificación
Amargasaurus y Dicraeosaurus junto con el saurópodo de cuello corto Brachytrachelopan se los colocan en la familia Dicraeosauridae. Los dicreosáuridos junto con los diplodócidos son incluidos dentro de Flagellicaudata.[3] [4]
La discusión comparativa de esta nueva especie sugiere que es más derivada que el Dicraeosaurus hansemanni de las últimas camas jurásicas de Tendaguru, en la bifurcación más alargada y más pronunciada de las vértebras presacras, y en los procesos más largos del basiterigoideo. La comparación de los géneros Dicraeosaurus - Amargasaurus con otros clados de saurópodos, particularmente con Diplodocidae, sugiere que correspondan a una familia sus los propia de saurópodos mas derivados con diferencias significativas a Diplodocidae.[2]
Allosaurus (gr. "reptil diferente") es un género de dinosaurios terópodos alosáuridos, que vivieron a finales del período Jurásico, hace aproximadamente 156 y 144 millones de años, en el Kimeridgiano y el Titoniano, en lo que hoy es Norteamérica y Europa. Se han encontrado varias especies aunque la más conocida en Allosaurus fragilis de Estados Unidos también se lo conoce de Portugal (A. europaeus) y posiblemente Tanzania, China y Siberia.
A diferencia de otros terópodos anteriores, los alosaurios eran comparativamente más grandes, de alrededor de 9 metros de largo, sin embargo compartían muchos de sus mismos rasgos anatómicos. Era un carnívoro bípedo con garras y patas masivas, balanceadas por su pesada cola. Su cráneo curvo provisto de dientes aserrados era característico de sus bajas crestas irregulares, sobre y delante de los ojos. A pesar de haber sido un temible cazador relativamente grande no pesaba tanto, lo mismo que un moderno rinoceronte indio.
Compartió el paisaje con varios saurópodos gigantescos como apatosaurio, diplodoco y camarasaurio así como con otros herbívoros tales como estegosaurio, driosaurio y camptosaurio, los cuales debieron haber sido su presa potencial. Fue el depredador grande más común en lo que hoy es Norteamérica, de hecho es famoso por ser el dinosaurio carnívoro más abundante y común en el registro fósil con cerca de 60 especímenes y unas cuantas docenas de ellos bien preservados. El gran número de alosaurios encontrado en un mismo sitio hace pensar que tenían una cierta vida en grupo.
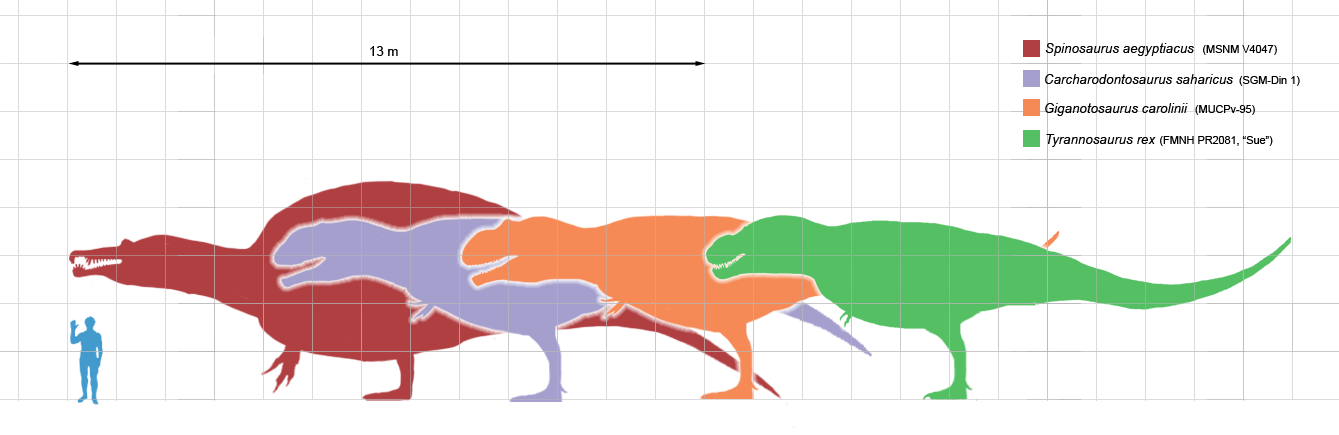
Los alosaurios presentaban un tamaño promedio de 8,5 metros de longitud y 3,5 metros de altura, exhibiendo un peso estimado de una tonelada.[1] Con un largo mayor conocido para un ejemplar de Allosaurus (AMNH 680) estimado en 9,7 metros,[2] y un peso estimado de 2,3 toneladas.[2] Como en todos los dinosaurios, el estimar su peso es un area de debate, desde la década de 1980 el rango de estimación a variado entre 1500 kilogramos, 1000 a 4000 kg, y 1010 para un adulto tipico, no el maximo peso posible.[3] John Foster, un especialista en la fauna de la Formación Morrison sugiere que , 1000 kg es un peso razonable para un adulto de A. fragilis, pero 700 kg es una estimación más cercana para los individuos medios según lo medido a través del fémur.[4]
En su trabajo de 1976 para Allosaurus, James Madsen menciona un rango de tamaño , según algunos huesos, con un máximo de 12 a 13 metros.[5] Esto se debe a que varios gigantes especímenes han sido históricamente referidos a Allosaurus, pero en realidad pertenecen a otro género. El cercano Saurophaganax (OMNH 1708) habría alcanzado quizás los 11 metros de largo,[2] siendo este único espécimen incluido dentro de Allosaurus como Allosaurus maximus, aunque recientes estudios apoyan la hipotesis de dos géneros separados.[6] Otro potencial Allosaurus, también asignado al género Epanterias (AMNH 5767), habría medido alrededor de 12 metros de largo.[2] El descubrimiento más reciente es un esqueleto parcial de la mina de Peterson en las rocas de Morrison de Nuevo México; este alosáuroide grande puede ser otro individuo de Saurophaganx.[7]
[editar] Cráneo
Cráneo de Allosaurus fragilis.
Cráneo de Allosaurus en el Museo de Historia Natural de San Diego.El cráneo y los dientes de Allosaurus tenia un tamaño modesto si lo comparamos con otros terópodos de su tamaño. A pesar de su tamaño, se ha encontrado una paradoja en su constitución: un cráneo fuerte con una mordida débil. Es por eso que los científicos creen que la forma de ataque de un Allosaurus era acechar a su presa y luego asestarle un fuerte golpe de mandíbulas moviendo su cabeza de arriba a abajo como un hacha. El paleontólogo Gregory S. Paul ha estimado un tamaño de 845 milímetros para el cráneo de un individuo de alrededor de 8 metros de largo.[8] A diferencia de otros terópodos, los alósauridos tenían un par de abultamientos, similares a "cuernos" embotados y dos líneas paralelas de pequeñas protuberancias óseas sobre el hueso nasal, al tope de la cabeza, sobre y delante de los ojos, teniendo gran variedad de formas y tamaños.[5] Estas ornamentaciones probablemente estuvieran cubierta de queratina lo que habría aumentado su tamaño. Se ha argumentado que poseían gran variedad de funciones, incluyendo que funcionaban para proteger del sol los ojos,[5] para exhibiciones, y para combates entre individuos de la misma especies[9] [8] aunque eran muy fragiles.[5] El cráneo también poseía un largo hocico y amplias fenestras que reducían el peso de la cabeza proporcionando áreas para la atadura de músculos y órganos sensoriales. Sus mandíbulas contenían cerca de 60 dientes afilados con forma de D en borde transversal, los cuales le hubieran ayudado a cazar presas y devorar carroña. El reborde óseo que tenían en la parte posterior de la cabeza era fuerte y rugoso, sirviendo de incerción muscular, también ha sido visto dentro de los tiranosáuridos.[8]
Dentro de los huesos lacrimales tenia unas depreciones para contener una glándula, similar a la glandula de sal.[10] Con los senos maxilares mejor desarrollados que en terópodos basales como Ceratosaurus y Marshosaurus, estando relacionado con el sentido del olfato, probablemente sosteniendo un órgano vomeronasal. El techo del cráneo era fino, quizás mejorar termorregulación para el cerebro.[5] En las mandíbula, los huesos del frente y la mitad trasera se articularon libremente, permitiendo que las quijadas se arqueen hacia fuera y aumentando la apertura de la boca.[11] El techo del cráneo y los frontales también tenían ese tipo de union.[5]
[editar] Esqueleto postcraneal
Replica del esqueleto de un alosaurio expuesto en el Museo de Historia Natural de San Diego.A pesar de que los miembros delanteros eran cortos en comparación a los miembros traseros, estos eran masivos y con garras parecidas a las del águila. El primer metacarpiano de cada "mano" es corto y robusto, y se encuentra girado lateralmente, lo que provocaba que su dedo correspondiente se dirigiese hacia los otros dos al cerrar la mano. El esqueleto del alosaurio, como otros terópodos, exhibía características de ave, así como la espoleta[12] [13] y vértebras huecas del cuello por sacos de aire, que usaba en la respiración.[14]
La caja costal era amplia, dándole un pecho con forma de barril, especialmente con respecto a los menos derivados terópodos como Ceratosaurus.[15] El Allosaurus tenía gastralias (costillas abdominales), pero no son comunes de encontrar,[5] debido al fosilizarse pobremente.[8] En un caso publicado, las gastralias muestran lesiones durante la vida.[16] La fúrcula estaba presente pero recién fue reconocida en 1996; en algunos casos se la confundida con gastralias.[17] [16] El ilion, el hueso principal de la cadera, era grande y fuerte, y el pubis tenía un saliente prominente que se pudo haber utilizado para la inserción de músculos y como apoyo para reclinar el cuerpo en la tierra. Madsen observó que alrededor mitad de los individuos de la mina de dinosaurios de Cleveland Lloyd, independiente del tamaño, los pubis no se había fundido al otro en los extremos de la saliente. Ha sugerido que esto es una característica sexual, donde las hembra carecieran de la fusión para poder poner los huevos mas fácilmente.[5] Esta propuesta no ha atraído la atención del medio científico.
Garra de A. fragilis.Los brazos de los alosaurios eran cortos en comparación a sus patas traseras, solo el 35% del largo de estas en un ejemplar adulto.[18] teniendo tres dedos por mano, terminados en unas garras, grandes y fuertes y curvadas.[5] Sus miembros superiores eran poderosos,[8] con el antebrazo mas corto que el brazo (una relación 1:1.2; entre húmero y ulna).[19] Este tenia una versión del hueso semilunar en el carpo[20] cosa encontrada en los terópodos mas derivados como en los maniraptores. Uno de esos dedos, el del medio, era más largo,[8] y se separaba de los otros.[19] Las patas no erán tan largas, o preparadas para la carrera como en los tiranosáuridos, y las garras de los dedos de los pies estaban menos desarrolladas y se parecían más a pezuñas que en los terópodos mas antiguos.[8] Cada pie tenia tres fuertes dedos que contactaban el piso y uno, como un garrón que Madsen sugirió se habría podido utilizase para agarrar en jóvenes.[5] También se ha interpretado que podía tener un remanente de un quinto metatarsal, que serviría como palanca entre el tendón de Aquiles y el pie.[21]
[editar] Historia
Esqueleto de un Allosaurus en el museo de la Universidad de Michigan.En 1869 los nativos de Middle Park, cerca de Granby, Colorado, hallaron una vértebra caudal incompleta, que fue entregada al geólogo Ferdinand Vandiveer Hayden, quien pensó que era la pezuña fosilizada de un caballo prehistórico. En 1870, el paleontólogo Joseph Leidy la estudió debidamente y asignó la vértebra al género Poekilopleuron, pero al notar la diferencia que esta vértebra tenía con las demás halladas hasta el momento, Leidy propuso que el fósil pertenecía a un nuevo género, al cual lo nombró Antrodemus,[22] sin embargo no fue un género formalmente descrito. Fue ya en 1877 que el célebre paleontólogo Othniel Charles Marsh le dio el nombre formal de Allosaurus al género, y A. fragilis a la especie tipo,[23] basado en mucho mejor material, incluyendo un esqueleto parcial, descubiertos en Garden Park, al norte de Cañon City, Colorado.
Al igual que muchos otros dinosaurios nombrados por Marsh, los restos de alosaurio fueron estudiados y descritos a toda velocidad durante la llamada Guerra de los Huesos que el autor sostuvo con su gran rival Edward Cope. Posteriores expediciones al oeste de Estados Unidos encontraron cada vez más restos en un proceso que dura hasta hoy convirtiendo a los alosaurios los dinosaurios mejor conocidos y uno de los carnívoros más grandes.
Uno de los hallazgos más significantes fue el descubrimiento de "Big Al"[24] (MOR 593) en 1991, un 95% completo, parcialmente articulado, espécimen subadulto que alcanzó los 10 metros de longitud. Tuvo 19 huesos rotos con señales de infección, lo cual probablemente contribuyó a su muerte.[25] Los fósiles fueron excavados cerca de Shell, Wyoming, por expedicionistas y paleontólogos del Museo de la Rocosas y del Museo Geológico de la Universidad de Wyoming. Este esqueleto fue inicialmente descubierto por un equipo suizo, dirigido por Kirby Siber. El mismo equipo luego excavó un segundo alosaurio, "Big Al Two".
[editar] Significado del nombre
Imagen en 3D de un Allosaurus.En 1877 se denominó el Allosaurus por Othniel Charles Marsh. El nombre genérico está compuesto de las palabras griegas αλλος (allos, "extraño") y σαυρος (sauros, "lagarto"),[26] en referencia a su vértebra diferente a la de todos los dinosaurios conocidos hasta ese momento y a su naturaleza reptiliana.[27] [28] El epíteto especifico fragilis proviene del Latín for "frágil", refirendoce a la ligera construcción del la vértebra.
[editar] Primeros descubrimientos
Allosaurus está basado en el holotipo YPM 1930, una pequeña colección de huesos que incluyen tres vértebras, fragmentos de costilla, un diente, un hueso del pie, a y lo más usado en posteriores discusiones, el eje del húmero derecho (miembro superior).[27] [28]
En sus ansias de encontrar muchos fósiles, Cope y Marsh no presentaban trabajos de sus descubrimientos (o, más comúnmente sucedió, esto se hizo por medio de sus subordinados). Por ejemplo, después del hallazgo de Benjamin Mudge del espécimen tipo de Allosaurus en Colorado, Marsh eligió concentrar sus esfuerzos en Wyoming; cuando reasumió el trabajo en Garden Park en 1883, M. P. Felch encontró un ejemplar completo de Allosaurus mas varios esqueletos parciales.[10] Además de esto, uno de los recolectores de Cope, H. F. Hubbell, encontró los especímenes de Como Bluff área de Wyoming en 1879, pero aparentemente no lo menciono por completo, y Cope nunca los desenterró. Fueron recogidos en 1903 (varios años después de la muerte de Cope), encontrándose uno de los esqueletos mas completos conocidos hasta hoy, y en 1908 el esqueleto, hoy catalogado como AMNH 5753, fue expuesto al publico.[29] Éste es el montaje bien conocido inclinado sobre un esqueleto parcial de Apatosaurus como si estuviese alimentándose, ilustrado como Charles R. Knight Es el primer montaje libre de un dinosaurio terópodo, e ilustrado y fotografiado a menudo, aunque nunca fuese descrito científicamente.[30]
Esqueleto de AMNH 5753 montado, alimentandoce de la cola de un Apatosaurus.
AMNH 5753 en la interpretación de la restauración de Charles R. Knight .La multiplicidad de nombres tempranos ha complicado estudios posteriores, con la situación aún más complicada por las descripciones concisas que proporcionaron Marsh y Cope. Por mucho tiempo autores como Samuel Wendell Williston sugirieron que existían muchos nombres para el mismo animal.[31] Por ejemplo, Williston apunta que en 1901 que Marsh nunca distiguió adecuadamente Allosaurus de Creosaurus.[32] La tentativa temprana más influyente de arreglar la situación fue producida por Charles W. Gilmore en 1920. Él llegó a la conclusión que la vértebra de la cola de ' ' Antrodemus' ' descrita por Leidy era indistinguible de las de Allosaurus , y así Antrodemus debe ser el nombre preferido porque como el más viejo nombre tenía prioridad.[19] Antrodemus se convirtió el nombre aceptado para este género por más de cincuenta años, hasta que James Madsen publicara los especímenes de Cleveland-Lloyd y concluyó que Allosaurus se debe utilizar ya que Antrodemus fue basado en el material con pobres características de diagnóstico y la pobre información del lugar de procedencia,(por ejemplo, formación geológica de Antrodemus es desconocida).[5] "Antrodemus" se ha utilizado informalmente para al distinguir entre el cráneo restaurado por Gilmore y el cráneo compuesto restaurado por Madsen. [33]
[editar] Descubrimientos de Cleveland-Lloyd
Una famosa "cama fósil" puede encontrarse en la Mina del Dinosaurio de Cleveland Lloyd, en Utah. Esta cama fósil contiene cerca de 10,000 huesos, sobre todo del alosaurio, entrelezado con restos de otros dinosaurios, como el estegosaurio y ceratosaurio. Es aún un misterio como los fósiles de tantos animales pueden hallarse en un solo lugar. El cociente de fósiles de animales carnívoros sobre los de herbívoros es normalmente muy pequeño. Descubrimientos como estos pueden indicar que el alosaurio cazaba en grupo, sin embargo esto es muy difícil de probar. Algo verdaderamente más posible es que el sitio de Cleveland Lloyd haya formado una "trampa de depredadores" (por ejemplo arenas movedizas), similar a Rancho La Brea, lo cual causó que un gran número de predadores hayan sido atraídos por la carne de herbívoros y luego capturados en un sedimento ineludible.[1]
Aunque aparecía de forma esporádica en trabajos desde 1927 la Mina de Dinosaurio de Cleveland-Lloyd en Condado de Emery, Utah y el lugar en sí mismo fue estudiado por William J. Stokes en 1945,[34] las mayores operaciones no comenzaron hasta 1960. En un esfuerzo cooperativo de unos 40 instituciones, cientos de huesos fueron recobrados entre 1960y 1965.[5] La mina es notable para el predominio de Allosaurus, con una excelente condición de los especímenes, y la carencia una explicación científica totalmente aceptada de cómo sucedió esta acumulación. La mayoria de los grandes terópodos son Allosaurus fragilis (por lo menos 46 A. fragilis , en un mínimo de 73 dinosaurios), y los fósiles se encontraron desarticulados y mezclados. Casi una docena de trabajos científicos se han escrito en tafonomía del sitio, presentando numerosas explicaciones contradictorias de su formación. Las sugerencias van desde que los animales habrían quedado atrapados en un pantano, en una trampa de fango, víctimas de una sequía muriendo alrededor de una charca que filtraba.[35] Sin importar la causa real, la gran cantidad de bien preservados Allosaurus han permitido que este género sea conocido detalladamente, haciéndolo entre los terópodos más conocidos. Los restos esqueléticos de la mina pertenecen a los individuos de casi todas las edades y tamaños, de 3 metros de largo[36] a 12 metros y lo disarticulado es una ventaja para describir los huesos, generalmente encontrados fundidos.[5]
[editar] Descubrimiento de 1980 hasta el presente
Representación de un Allosaurus.El periodo desde el trabajo de Madsen ha sido marcado por la gran cantidad de trabajo que hicieron foco en la vida del Allosaurus (sobre la paleobiolgía y paleoecología). Tales estudios han cubierto asuntos incluyendo la variación esquelética ,[37] desarrollo,[38] [39] conformación del cráneo,[40] métodos de caza,[41] el cerebro,[42] y la posibilidad de vida gregaria y del cuidado de las crías por los padres.[43] El nuevo análisis de material viejo (particularmente de los especímenes más grandes),[8] [44] nuevos descubrimientos en Portugal,[45] varios nuevos ejemplares completos encontrados[46] [16] [47] cosas que hicieron aumentar el conocimiento de este dinosaurio.
[editar] "Big Al"
Uno de los más significativo hallazgo de Allosaurus encontrados en 1991 fue el descubrimiento de "Big Al" (MOR 693), un espécimen 95% completo y parcialmente articulado, que midió alrededor de 10 metros de largo . MOR 693 fue excavado cerca de Shell, Wyoming, por una equipo del Museo de las Rocallosas y la Universidad de Wyoming.[48] Este esqueleto fue descubierto por un equipo suizo, liderado por Kirby Siber. Este mismo equipo excavó un segundo espécimen deAllosaurus, "Big Al Dos", el cual es el mejor esqueleto preservado de su clase hasta la fecha.[47]
Lo completo, la preservación, y la importancia científica de este esqueleto dieron el "Big Al", su nombre, el individuo en sí mismo estaba debajo del tamaño medio para Allosaurus fragilis[48] y era un subadulto que habría alcanzado un 87% del tamaño adulto.[49] El espécimen lo describió Breithaupt en 1996.[46] Diecinueve de sus huesos estaban quebrados o mostraban signos de infección, que pudieron haber contribuido la muerte de "Big Al". Las patologías de los huesos incluían cinco costillas, cinco vértebras, y cuatro huesos de los pies; varios huesos dañados mostrando osteomielitis, una infección del hueso. Un problema particular para el animal vivo era infección y trauma al pie derecho que probablemente afectaba el movimiento y pudo también haber predispuesto el otro pie a lesión debido a un cambio en el paso.[49]
[editar] Clasificación
Allosaurus es un miembro de la familia Allosauridae de grandes terópodos dentro del gran grupo Carnosauria. El nombre de la familia fue crado a partir de este género en 1878 por Othniel Charles Marsh,[50] pero el termino cayo en desuso hasta los años de 1970 en favor de Megalosauridae, otra familia de grandes terópodos que se convertiría en un taxón cajón de sastre. Esto junto con el uso de Antrodemus por Allosaurus durante el mismo periodo, es un punto que es necesario recordar al revisar la información sobre Allosaurus en publicaciones anteriores al trabajo de James Madsen en 1976. La mayor parte de la publicaciones que usaron el nombre de Megalosauridae en vez de Allosauridae incluye trabajos de Gilmore, 1920,[19] von Huene, 1926,[51] Romer, 1956 y 1966,[52] [53] Steel, 1970,[54] y Walker, 1964.[55]
Luego del influyente trabajo de Madsen, Allosauridae se convirtió en la familia preferida para clasificarlo, pero esta no se encontraba fuertemente definida. Durante ese periodo se usó una gran variedad de grandes terópodos para definir Allosauridae, usualmente los más grandes y mejor conocidos que los megalosáuridos. Los típicos terópodos con los que se relaciono a Allosaurus incluyen a Indosaurus (un abelisáurido), Piatnitzkysaurus (un tetanuro basal), Piveteausaurus (un megalosáurido), Yangchuanosaurus (un sinraptórido),[56] Acrocanthosaurus (un carcarodontosáurido), Chilantaisaurus (un espinosáurido), Compsosuchus (un abelisáurido), Stokesosaurus (un tiranosauroide basal), y Szechuanosaurus (un sinraptórido).[57] Con el moderno conocimiento de la diversidad de los terópodos y las ventajas de la cládistica aplicada en el estudio de las relaciones entre las especies, ninguno de esos terópodos estoy considerado un alosáurido, aunque varios de estos como Acrocanthosaurus y Yangchuanosaurus, son miembros de familias cercanas.[14]
Esqueleto del Allosaurus en el museo de Canterbury, Christchurch, Nueva Zelanda.Allosaurus es el género tipo de la familia Allosauridae, la cual fue también nombrada por Marsh en 1878. En la taxonomía filogenética, el grupo Allosauridae es usualmente definido como "todos los carnosaurios más cercanamente relacionados con Allosaurus que con el Sinraptor o Carcharodontosaurus". Cuatro géneros han sido formalmente descritos los cuales son el Allosaurus y sus relativos más cercanos, otros dos géneros, "Madsenius"[58] y "Wyomingraptor",[59] no han sido nombrados formalmente y tampoco se conocen por muchos restos.
Allosauridae es una de las tres familias de Carnosauria; las otras dos son Carcharodontosauridae y Sinraptoridae.[14] Allosauridae fue en un tiempo propuesto como ancestro de la familia Tyrannosauridae (lo cuál los haría parafiléticos), con un ejemplo reciente en Gregory S. Paul's Predatory Dinosaurs of the World,[60] pero esto ha sido rechazado ya que los tiranosáuroideos han sido identificados como un miembro de Coelurosauria, un grupo totalmente separado de terópodos.[61] Allosauridae es la más pequeña familia dentro de Carnosauria, con solo Saurophaganax y un alosáuroide Francés sin describir aceptados como géneros validos aparte de Allosaurus en las revisiones más recientes.[14] El otro género, Epanterias, es un potencial valido, el cual junto con Saurophaganax es nombrado como ejemplo de Allosaurus gigantes.[8] Las ultimas revisiones aceptan mantener a Saurophaganax e incluyen a Epanterias con Allosaurus.[3] [14]
[editar] Especies de Allosaurus
Allosaurus se uso como sinónimo probable de los géneros Antrodemus , Creosaurus , Epanterias , y Labrosaurus. La mayor parte de las especies que se ven como sinónimos de A. fragilis, o han sido apartados del género, debido a estar basado en material oscuro y escaso. Una excepción es Labrosaurus ferox, nombrado en 1884 por Marsh para una mandíbula inferior parcial de formada extraña, con un boquete prominente en la fila del diente en el extremo, y una sección posterior ampliada y girada grandemente abajo.[62] Investigadores posteriores sugirieron que fuera el patología hueso, mostrando una lesión en el animal vivo, y esa forma inusual de la parte posterior del hueso era debido a la reconstrucción del yeso.[63] Siendo hoy considerado un ejemplo de A. fragilis.[14]
Cráneo de la especie sin nombre de Allosaurus procedente de la Formación de Morrison.
Comparación de tamaños entre el ser humano y diferentes especies de Allosaurus.Se han identificado varias especies de Allosaurus a lo largo de la historia, sin embargo muchas de ellas han resultado ser sinónimos de la especie tipo Allosaurus fragilis o transladadas a otros géneros, con la posible excepción del espécimen de Epanterias amplexus (AMNH 5767), el cual mide 12 metros de longitud y podría ser un Allosaurus completamente adulto o un saurofaganax; el espécimen más grande de A. fragilis (AMNH 680) mide 9,7 metros de longitud, y al este ser adulto la mayoría de científicos y paleontólogos ahora consideran al Allosaurus con una longitud promedio de 8 y 9 metros. El género cercanamente relacionado Saurophaganax (OMNH 1708) alcanzó los 11 metros, y es a veces incluido en el género Allosaurus como A. maximus, pero recientes estudios apoyan la idea de que pertenece a un género separado.[64] Otras especies que se creyeron pertenecer al género Allosaurus también fueron desplazadas a otros géneros.
Un Allosaurus alimentándose.La especie más extendida fue A.fragilis que habitó en EEUU y en Portugal había una especie muy relacionada, ya que en aquel entonces estaban conectados.[65] Normalmente los holotipos de estas especies válidas son esqueletos parciales e incompletos, pero existe la excepción de una especie aún no nombrada, que se conoce por varios restos fósiles y esqueletos casi completos.[66] El espécimen de "Big Al" (MOR 693) pertenece a esta especie y al tener un 95% de sus restos recobrados da una gran idea de como pudieron haber sido, sin embargo, generalmente no se han examinado muy bien los demás especímenes de esta especie por lo que permanece sin nombre.
A. amplexus, A. atrox, A. fragilis, "A. jimmadseni" y A. maximus son todas conocidas del intervalo Kimeridgiano–Titoniano en el Jurásico superior de la Formación Morrison de los Estados Unidos, aparecdiendo a través de los estados de Colorado, Montana, New Mexico, Oklahoma, Dakota del Sur, Utah y Wyoming. A. fragilis es sin duda la más común, con alrededor de sesenta especímenes conocidos.[14] La discusión ocurre desde los años 80 con respecto a la posibilidad que haya dos especies comunes en la Formación Morrison de Allosaurus , con la segunda conocida como ' ' A. atrox' ';[8] [67] Un trabajo reciente ha seguido una interpretación de "un especie",[14] con las diferencias vistas en el material de la Formación Morrison atribuido a la variación individual.[68] [69] A. europaeus en sedimentos del Kimeridgiano del Miembro Porto Novo de la Formación Lourinhã,[70] pero podría ser un A. fragilis.[71] A. tendagurensis fue encontrado en rocas del Kimeridgiano de Tendaguru, en Mtwara, Tanzania.[72] Aunque la revisión más reciente lo aceptara tentativo como especie válida de Allosaurus , puede ser un tetanuro más básico,[73] o un simple terópodo dudóso.[1] Aunque sea oscuro, fue un terópodo grande, posiblemente alrededor de 10 metros de largo y 2,5 toneladas de peso,[2] del cual solo se ha encontrado una tibia parcial.[74] También se incluye al "A. robustus" de Australia,[75] [76] el cual actualmente se concluyó que era un carnosaurio basal por su astrágalo (hueso del tobillo) muy similar al del fukuirráptor. Una vértebra caudal fragmentaria de otra posible especie ha sido hallada en la provincia de Shanxi, China,[77] y seis dientes que datan del Jurásico medio y superior han sido recobrados de la Formación de Djaskoian, Rusia.,[78] [79] y en Suiza,[1]
Todavía no esta claro el número de especies conocidas de Allosaurus. Al menos siete especies han sido consideradas potenciales validas desde 1988 (A. amplexus,[8] A. atrox,[8] A. europaeus,[70] la especie tipo A. fragilis,[14] , la todavía no descrita formalmente "A. jimmadseni",[6] A. maximus,[37] y A. tendagurensis[14] ), aunque generalmente solamente una fracción de estas sea considera válida en un momento dado. Además, hay por lo menos diez especies dudosas o sin describir que se han asignado a Allosaurus, junto con la especie que pertenece a los géneros hoy inválidos de Allosaurus. En la revisión más reciente de tetanuros basales, solamente A. fragilis (' incluyendo; ' A. amplexus' ' y ' ' A. atrox' ' como sinónimos), A "Jimmadseni".; (como especie sin nombre), y A. tendagurensis fueron aceptados como especie potencialmente válida, con A. europaeus no todavía propuesta y A. maximus asignado a Saurophaganax.[14]
Mamenchisaurus (zh. "reptil de Mamenchi") es un género de dinosaurios saurópodos mamenquisáuridos, que vivieron a finales del período Jurásico, hace aproximadamente 156 millones de años, en el Oxfordiano, en lo que hoy es Asia. Mamenchisaurus significa 'lagarto de Mamenchi', del Pinyin mǎ (马 'horse') y mén (门 ' puerta'), mientras que chi es la transliteración de xī (溪 'corriente' o ' arroyo'), combinadas con el sufijo griego -saurus ('lagarto'). Fue nombrado por el lugar en donde su primer fósil fue encontrado, el sitio de construcción, al lado del cruce en ferry de Mǎmíngxī (马鸣溪) por el Río Jinsha (金沙江, la corriente principal más al oeste de la cabecera del Río Yangtze), cerca de Yibin (宜宾) en la provincia de Sicuani de China.
Sin embargo, debido a una confusión acentual de Young, el nombre del lugar Mǎmíngxī (马鸣溪 'horse-neighing brook') fue cambiado por Mǎménxī (马门溪 'horse-gate brook').[1] El hecho es que el fósil de Mamenchisaurus fue excavado de un emplazamiento de la obra llevado a cabo por Young por lo que nombro a la especie tipo Mamenchisaurus constructus.
Los mamenquisaurios debían medir entre 25 metros de largo en las especies más grandes a 16 las pequeñas. El primer esqueleto encontrado medía 18,5 metros, pero tenía la cola incompleta, de los cuales 9 correspondían a las 19 vértebras huecas de que se componía el cuello, uno de los más largos conocido, y pesaba 27 toneladas. Como sus parientes, los diplodocos, los mamenquisaurios tenían las patas traseras más altas que las delanteras. Al parecer viajaba en manadas, posiblemente cuando su alimento escaseaba. Era herbívoro y se alimentaba del follaje de los árboles.
Ilustración de un Mamenchisaurus.Las vértebras cervicales están alargadas, que junto con el número creciente de las mismas, producen el cuello más largo en relacición al cuerpo conocido para un animal terrestre. Algunos individuos pudieron haber tenido cuellos de 15 metros de largo. Con todo el cuello era comparativamente ligero, algunas áreas del hueso tenían el grosor de la cáscara de un huevo. Cada las vértebras cervicales tenían dos costillas cilíndricas para fortalecer el cuello, como en otros saurópodos. Algunas características de la espina dorsal son evocadoras al Diplodocus. El número total de caudales y si los extremos de cola formaban un látigo no se conocen. Los cheurones comienzan a bifurcarse en la caudal 12, y los cheurones posteriores muestran que la extensión longitudinalmente está desarrollada más que en cualquier otro género con excepción de Diplodocus en sí mismo. Los fragmentos del cráneo encontraron en la década de los 1980 sugieren que Mamenschisaurus tenía un hocico más alto y cuadrado que Diplodocus, y dientes eran más robustos y con forma de una cuchara.bwdfg3fdrg3f
[editar] Historia
Restauración de Mamenchisaurus youngi.Los restos fósiles de Mamenchisaurus fueron descubiertos en 1952 en Sichuan, Formación Shangshaximiao en el centro-sur de China durante la construcción de una carretera. El fósil, un esqueleto parcial postcraneal fue estudiado, y nombrado en 1954, por el renombrado del paleontólogo chino Profesor C. C. Young.[2] El primer especímen encontrado (el holotipo) media alrededor de 22 metros de los cuales la mitad aproximadamente pertenecia la cuello, cuál le hizo el cuello conocido más largo de cualquier animal en ese entonces. 19 vértebras fueron descubiertas (otro record), con sus costillas cervicales. Otros especímenes fueron también encontrados en Formación Hantong, Gansu.
Mamenchisaurus, Museo Field de Historia Natural.En 1972, una segunda especie de Mamenchisaurus fue descubierta en la Formación Shangshaximiao. Conocida por los restos de 4 individuos, M. hochuanensis, tiene un cuello de alrededor de 9,5 metros de largo, un largo total de 22 metros y 3,5 metros a la cruz y 48 toneladas de peso.[3] Hasta 1994, cuando el Sauroposeidon que fuera descubierto en Estados Unidos, con un cuello estimado de entre 10,5 a 11,5 metros fueron los cuellos más largos conocidos, aunque su cuello no excedió el del conocido previamente Supersaurus, con un largo de cuello de entre 13–14 metros.[4]
Mamenchisaurus jingyanesi del.Museo de Beijing, en la gira por el Museo de Ciencias de Miami.Una tercer especie, M. sinocanadorum fue descrita en 1993, se conoce por restos provenientes del Bajio Junggar, Formación Shishugou,[5] esta especie poseía las costillas cervicales más larga descritas en un dinosaurio saurópodo, midiendo 4,1 metros. Estas son más largas que las mayores costillas cervicales del Sauroposeidon que median 3,42 metros.[6] La cuarta especie M. youngi fue descrita por Pi, Ouyang & Ye en 1996 como M. youngi fue desenterrado en el Condado de Xinmin, cerca de la Ciudad de Zigong en Sichuan, China, en 1989, en sedimentos de la Formación Shangshaximiao.[7] El especimen fósil mide 16 metros de largo con un cuello de 6,5 metros, es relativamente más pequeña que las otras especies de Mamenchisaurus. El nombre especifico fue puesto en honor a C.C. Young. Una quinta especie, descrita por He, Yang, Cai, Li enLiu Z. en 1996 recibio el nombre de M. anyuensis y mide más de 21 metros de largo del Bajio Sichuan.[8] La ultima especie es M. jingyanensis descrita por Zhang, Li y Zeng en 1998, del Bajio Sichuan, Formación Suining y Formación Penglaizhen.[9]
Gallimimus (gr. "imitador de gallina") es un género representado por una única especie de dinosaurio terópodo ornitomímido, que vivió a finales del período Cretácico, hace aproximadamente 83 y 65 millones de años en el Campaniano y el Mastrichtiano, en lo que hoy es Asia. Fue hallado en la Formación Nemegt, en Mongolia. Debido a su constitución ligera y sus largas patas traseras, este dinosaurio estaba adaptado para la carrera. Sin duda fue esa su gran braza para escapar de los depredadores. Su aspecto recuerda al de un avestruz dado su largo cuello y su pico sin dientes; no obstante, no hay constancia de que tuviera alas o plumas. Su larga cola le servía para mantener el equilibrio al avanzar. Parte de su fama se la debe a su aparición en la saga de novelas de Michael Crichton El Mundo Perdido, y de la primera entrega de la saga de películas Parque Jurásico dirigida por Steven Spielberg.Gallimimus era en apariencia similar a los actuales avestruces, cabeza pequeña, grandes ojos, cuello largo, miembros anteriores pequeños, fuertes patas para correr y una larga cola. Un carácter de diagnóstico de Gallimimus son sus manos cortas en comparación al húmero, cuando se lo compara con otros ornitomímidos. Sus patas delanteras eran relativamente cortas comparadas con las traseras, y estaban rematados con tres garras en cada mano. A pesar de estar afiladas, no está claro que tuvieran mucha utilidad. Algunos expertos han dado utilidad a las garras del Gallimimus aventurando que pudieron servirle para escarbar en la tierra y conseguir así huevos con los que alimentarse. El cociente del antebrazo con el brazo alcanza el 66% en Gallimimus, mientras que en las formas americanas varia del 70% a incluso el 91% en Dromiceiomimus. Otra diferencias con sus parientes de Norteamérica se ven en las comparciones de las longitudes de los huesos. En Gallimimus el cociente de las vértebras dorsales con las caudales del 85% es mayor que en otros géneros, en Struthiomimus, por ejemplo, este cociente es el aproximadamente 76%. Las sacrales en Gallimimus también se distinguen por su gran alargamiento concerniente a la longitud del fémur. Además de esto, en la forma mongol los arcos neurales dorsales de las vértebras sacras no se funden totalmente incluso en especímenes maduros, en contraste con ésos observados en Ornithomimus y Struthiomimus. La cola era utilizada como contrapeso. Los ojos estaban situados a los lados de su cabeza, significando que no poseía la visión binocular. Como la mayoría de los pájaros modernos, tenía huesos huecos. Gallimimus tenía un número de adaptaciones que sugieren buena capacidad para correr, miembros largos, con una tibia y metatarso largos y los dedos del pie cortos. Un pico fosilizado presente en un cráneo de Gallimimus estaba bordeado de un pico que se ha interpretado como parte de un mecanismo de filtrado como los de los patos. Sin embargo, picos similares se ven en tortugas de mar herbívoras y los ornitomímidos eran relativamente comunes en ambientes con estaciones secas, donde no había alimentación para filtrar. Parece más probable que Gallimimus fuese un omnívoro, usando su pico en las plantas y la captura de pequeños animales. Sobre el Gallimimus se ha especulado sobre su dieta, sugiriendo que pudo haberse alimentado de insectos y de pequeños gusanos y pudo complementar su dieta insectívora con algunas plantas y huevos. Fue uno de los ornitomímidos más grandes, llegó a medir hasta 6 metros y pesar 440 kilogramos.
[editar] Historia
Esqueleto de gallimimo.[editar] Descubrimiento
Los primeros restos fósiles del gallimimo fueron descubiertos a principios de 1970, en el Desierto de Gobi. Ya para 1972, los paleontólogos Rinchen Barsbold, Halszka Osmólska, y Ewa Roniewicz nombraron al género. La única especie conocida y tipo Gallimimus bullatus. Una supuesta segunda especie, "Gallimimus mongoliensis", nunca fue formalmente referida al género. Un reciente reanalisis del esqueleto casi completo de "G. mongoliensis" concluyo que no pertenece a Gallimimus pero represente un nuevo, todavía no nombrado género de ornitomímido.[1]
Nuevos fósiles descubiertos por Peter J. Makovicky en el Desierto de Gobi en el 2001, indican que el gallimimo poseía un pico que filtraba de los charcos animales diminutos y agua a través de unas ranuras o lamelas que tienen en el pico y la lengua como los flamencos actuales.
El primer espécimen fue descrito y nombrado en 1840 a partir de un fragmento de mandíbula descubierto al suroeste de Inglaterra, en la localidad de Bristol. Otros descubrimientos iniciales incluyen dientes, mandíbulas, vértebras, costillas y restos de extremidades traseras y delanteras. Desde su hallazgo inicial, se han descubierto al sur de Inglaterra y Gales muchos más esqueletos parciales y cráneos de tecodontosaurio, casi todos en antiguas cuevas o fisuras. Al parecer algunos de estos pequeños dinosaurios cayeron en grietas rocosas al pasar por la superficie caliza del Triásico Superior. Con el tiempo las grietas se llenaron de lodo y arena y los cuerpos de los animales quedaron atrapados y preservados en la roca. Los restos fósiles de Thecodontosaurus, y otro material relacionado, fueron destruidos en 1940, en los bombardeos de la Segunda Guerra Mundial. Sin embargo, más restos han sido encontrados en un número de localidades, incluyendo nuevamente Bristol. Algo de este nuevo material pertenece a un espécimen juvenil que se creyó corresponder a una especie distinta, Thecodontosaurus caducus,[1] sin embargo recientemente al haberse identificado diferencias en las vértebras cervicales, cráneo, y cresta deltopectal, se asignó a esta especie al género Pantydraco, conociéndose ahora como P. caducus.[2] El dinosaurio australiano Agrosaurus macgillivrayi es probablemente sinónimo con el Thecodontosaurus antiquus.
[editar] Clasificación
Originalmente fue incluido en el clado Prosauropoda[3] pero más recientemente se ha sugerido que el tecodontosaurio y sus relativos fueron prioridad de la fractura Prosaurópodo-Saurópodo.[4] Nuevas reconstrucciones muestran que su cuello es proporcionalmente más corto que en sauropodomorfos antiguos más avanzados. Aunque no es actualmente el miembro más antiguo del grupo,ese honor pertenece a unos ejemplares de sauropodomorfos aún no nombrados procedentes de Madagascar,[5] Thecodontosaurus es el representante más primitivo que mejor se conoce de dinosaurios sauropodomorfos.
A diferencia de otros saurópodos, Brachiosaurus tenía una constitución similar a la de las actuales jirafas, con largas patas delanteras y un largo cuello, el cual, probablemente, utilizaba para alimentarse de las copas de los árboles. En cuanto a sus extremidades, el primer dedo de sus patas delanteras y los tres primeros de sus patas traseras tenían garras.
Brachiosaurus alimentándose de las gimnospermas.Se estima que pesaba de 35 a 60 toneladas y que podía llegar a medir 13 metros de altura y 25 metros de largo. Además, tenía dientes en forma de espátula y fosas nasales en la parte superior de su cabeza; esto último podría llegar a indicar que poseía un buen sentido del olfato. También tenía orificios en su cráneo para reducir el peso.
Se solía creer que el Brachiosaurus utilizaba sus fosas nasales para bucear, y que pasaba la mayor parte del tiempo sumergido en el agua para compensar su gran masa corporal. Sin embargo, la teoría aceptada actualmente sostiene que era un animal terrestre por naturaleza, algunos estudios han indicado que la presión del agua habría sido demasiado grande para que pudiera respirar estando sumergido. Además, sus patas eran muy angostas y al sumergirse en el agua se habría hundido en el lodo.
Por muchas décadas, Brachiosaurus fue el dinosaurio más grande conocido. Se ha descubierto desde entonces que un número de titanosaurianos gigantes (Argentinosaurus, por ejemplo) que sobrepasaron a Brachiosaurus en términos de masa. Más recientemente, se ha descubierto otro braquiosáurido, Sauroposeidon, que de acuerdo con la evidencia fósil incompleta, probablemente también haya sobrepasado a Brachiosaurus.
Brachiosaurus es a menudo considerado como el dinosaurio más grande conocido por un esqueleto fosilizado relativamente completo. Sin embargo, los especímenes más completos, incluyendo el Brachiosaurus del Museo de Ciencias Naturales de Berlín ("Museo Humbodlt"), excavado en África, son miembros de la especie B. brancai, y que algunos científicos la consideran parte de un género separado, Giraffatitan. El material del holotipo de la especie tipo, B. altithorax incluye una secuencia de siete vértebras dorsales posteriores, sacro, vértebra caudal, coracoides próximal, húmero, fémur y las costillas, con lo cual se puede estimar el tamaño. De acuerdo con un esqueleto compuesto completo, Brachiosaurus alcanzó 25 metros de longitud y podía probablemente levantar su cabeza cerca de 13 metros sobre el nivel del suelo. Material fragmentario de especímenes más grandes indica que podría crecer el 15% más de largo que esto. Tal material incluye un peroné aislado (HMN XV2) de 1340 mm de largo y el escapulo-coracoides de braquiosáurido referido a "Ultrasauros".
Las estimaciones históricas del peso de Brachiosaurus han variado entre las 15 toneladas (Russell et al., 1980) y las 78 toneladas.[1] Sin embargo estas estimaciones extremas ahora se consideran inverosímiles; la de Russell et al. fue basada en la alometría del miembro-hueso en vez de un modelo del cuerpo, y Colbert se basó en un modelo anticuado y gordo. Estimaciones más recientes basadas en los modelos reconstruidos a partir de la osteología y de la musculatura deducida están en el rango de 32[2] a 37 toneladas (Christiansen, 1997). Los especímenes más largos a los que se hizo alusión anteriormente habrían rondado entre 48 y 56 toneladas.
[editar] Cráneo
Cráneo Brachiosaurus brancai, Naturkundemuseum Berlin.Brachiosaurus ha sido caracterizado tradicionalmente por su cráneo alto con cresta distintiva, aunque esto pudo haber sido solo una característica de B. brancai. Otro cráneo completo de Brachiosaurus conocido, es el que usara Marsh en las primeras reconstrucciones de Brontosaurus. Carpenter y Tidwell estudiaron este cráneo 1998 y encontraron que pertenecía a una especie norteamericana de Brachiosaurus. Este cráneo se parece más al de Camarasaurus, con forma de caja, que al de B. brancai con la cresta sobre la nariz.
[editar] Cerebro
Como otros saurópodos, Brachiosaurus tenia un cerebro relativamente pequeño, incluso cuando se considera su enorme tamaño de cuerpo. Un estudio de 2009 calculaba su cociente cerebro/cuerpo, un cálculo aproximado de la inteligencia posible, entre un 0,62 a 0,79, dependiendo de la estimación del tamaño utilizado. Brachiosaurus es también similar a otros saurópodos en tener una ampliación del canal espinal sobre las caderas, a que algunas de las más viejas fuentes refirieron engañosamente como "segundo cerebro".[3]
[editar] Historia
Comparación de tamaño entre el Brachiosaurus y un humanoEl primer braquiosauro fue descubierto en el año 1900 por Elmer S. Riggs en el Río del Gran Cañón del oeste de Colorado, en los Estados Unidos. Bautizó a este nuevo género en 1903 refiriéndose a sus largas patas delanteras.
Desde el año 1909, Werner Janensch encontró numerosos especímenes nuevos —asignados a la especie Brachiosaurus brancai—, en Tendaguru (Tanzania), los primeros del continente africano, incluyendo algunos esqueletos casi completos, los cuales fueron usados para la reconstrucción de Berlín. Actualmente pertenece al género Giraffatitan.
[editar] Clasificación
Braquisaurus es un gran Sauropoda Macronaria de la familia Brachiosauridae, a la que le da el nombre.
[editar] Especies
Hay una especie conocida de Brachiosaurus y posiblemente otra más:
B. altithorax (Riggs, 1903): Se conoció gracias al descubrimiento de dos esqueletos parcialmente completos en Colorado y Utah, en los Estados Unidos. Vivió hace unos 145-150 millones de años durante el período Jurásico tardío.
Brachiosaurus altithorax.B. nougaredi (de Lapparent, 1960): Si bien es posible que no se trate de una especie diferente, se conoció gracias al descubrimiento de unos huesos fusionados ubicados sobre la cadera (sacro) y partes de un antebrazo en Wargla, Argelia (África). Vivió hace unos 100-110 millones de años, a mediados del período Cretáceo.
[editar] Especies extraídas del género
Además de la recientemente separada Giraffatitan, B. alataiensis, descrita por de Lapparent y Zbyszewski en 1957 ha sido referida a un nuevo género Lusotitan (Antunes & Mateus 2003). Es conocido por una columna vertebral y partes de la cadera y miembros que se han recobrados en Estremadura, Portugal. Vivió a finales del Jurásico hace 150 millones de años en el Kimeridgiano.[4]
[editar] Paleobiología
Si los braquiosaurios hubiesen sido endotérmicos (sangre caliente), debieron haber tardado 10 años para alcanzar su madurez, pero si hubiese sido poiquilotermo (sangre fría) hubiesen requerido más de 100 años. Como animal de sangre caliente, las demandas energéticas diarias de un braquiosaurio habrían sido enormes, necesitando probablemente comer más que 182 kilogramos de alimento por día. En cambio si el braquiosaurio era completamente de sangre fría o una pasiva gigantotermia, habría necesitado mucho menos alimento para cubrir sus necesidades energéticas diarias. Algunos científicos han propuesto que los dinosaurios grandes como el braquiosaurio se mantenían calientes por su enorme tamaño (gigantotermia).[5]
[editar] Paleoecología
Hueso de la pata delantera de Brachiosaurus.Los braquiosaurios eran uno de los más grandes dinosaurios del Jurásico, viviendo en las praderas llenas de helechos, bennetites y equisetos, moviéndose en vastos bosques de coníferas y arboledas de cícadas, helechos con semilla y ginkgos. Algunos géneros de dinosaurios contemporáneos incluyen a Stegosaurus, Dryosaurus, Apatosaurus y Diplodocus. Mientras que se especula que los grupos de braquiosaurios se movían en manadas, los individuos completamente desarrollados tenían poco que temer incluso de los depredadores más grandes de su tiempo, como Allosaurus y Torvosaurus, a causa de su tamaño menor.
Las narinas de los braquiosaurios, como las enormes aberturas nasales correspondientes en su cráneo, estaban situadas en lo más alto de la cabeza. Décadas atrás, los científicos supusieron que el animal utilizó sus ventanas nasales como un tubo respirador, pasando la mayor parte de su tiempo sumergido en agua para sustentar su gran masa corporal. La opinión de consenso actual, sin embargo, es que los braquiosaurios eran completamente terrestres. Los estudios han demostrado que la presión del agua habría evitado que el animal respirara con eficacia mientras que estaba sumergido y que sus pies eran demasiado estrechos para el uso acuático eficiente. Además, los nuevos estudios de Larry Witmer (2001) demuestran que, mientras que las aberturas nasales en el cráneo estaban puestas arriba sobre los ojos, las ventanas de la nariz todavía habrían estado cercanas a la extremidad del hocico, este estudio también presta apoyo a la idea que las "altas crestas" de los braquiosaurios soportaban una cierta clase de compartimiento de resonancia carnosa.
[editar] En la cultura popular
Animatrónico de Brachiosaurus con las nárinas en el frente del hocico, en la exhibición Dino Jaws exhibition - museo de Historia Natural de Londres.Brachiosaurus es uno de los dinosaurios más conocidos entre los paleontólogos y el público en general. Como tal, el género ha aparecido en muchas películas y programas de televisión, especialmente Parque Jurásico, Parque Jurásico III y Walking with Dinosaurs.
Un asteroide, en el cinturón de asteroides, 1991 GX7, fue llamado 9954 Brachiosaurus en honor al género.[6] [7]
[editar] En los museos
Una réplica del esqueleto de un Brachiosaurus está montado desde enero de 2000 en la sala B de la Terminal Uno de United Airlines en el Aeropuerto Internacional O'Hare de Chicago (Estados Unidos), cortesía del Museo Field de Historia Natural de Chicago. Otra, idéntica, pero esta vez en bronce en el exterior del propio museo
[editar] Descripción
El mayor espécimen conocido de Deinonychus alcanzó los 3,4 metros de largo, con un cráneo de alrededor de 410 milímetros, una altura de cadera de 87 centímetros y un peso máximo de 73 kilogramos.[2] Poseía una gran cabeza, con alrededor sesenta dientes curvados hacia atrás, con forma de cuchillos. Los estudios del cráneo han progresado mucho durante las últimas décadas. Ostrom lo reconstruyó a partir de ejemplares parciales preservados imperfectamente, describiéndolos como triangulares, amplios, y bastante similares a los del Allosaurus. Material adicional del cráneo de Deinonychus y géneros estrechamente vinculados encontrados con una buena preservación tridimensional[3] muestran que el paladar era más estrecho que lo que pensó Ostrom, haciendo el hocico más alargado, mientras que el yugal, flanqueado por aberturas, daba una mayor visión estereoscópica. El cráneo de Deinonychus era diferente en la forma al de Velociraptor , con un techo del cráneo como el de Dromaeosaurus , y no tenía los nasales deprimidos de Velociraptor.[3] El cráneo y la mandíbula inferior tenían ventanas para reducir el peso del cráneo. En Deinonychus la ventana anteorbital, una abertura del cráneo entre el ojo y las narinas, eran particularmente grandes.[3]
Su cuello arqueado en forma de S, extremidades largas y fornidas y el segundo dedo del pie provisto de la extraordinaria y terrible garra falciforme, que el animal alzaba del suelo para conservar su aguda punta, y cuya función no era otra sino que desgarrar y destripar. El primer dedo estaba atrofiado, el quinto ausente y el tercero y cuarto se usaban para caminar. En las garras delanteras había sólo tres dedos igual de fuertes y coronados por garras curvas del mismo tamaño, el primer dígito era el más corto y el segundo era el más largo.. Cuando no se usaban para aferrar a la presa, estas garras se recogían bajo el brazo, de forma similar a como un ave recoge sus alas.
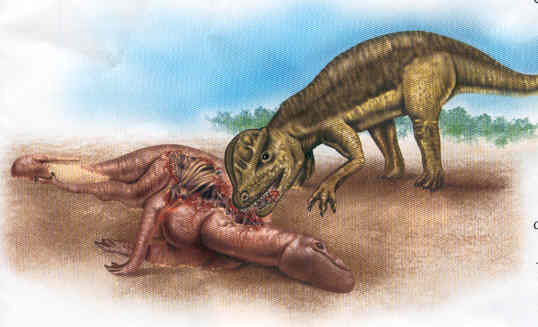
Comparación de tamaño del deinonico con un ser humano.Yacimientos con cientos de individuos fosilizados y esqueletos de otros dinosaurios con dientes de deinonico de distinto tamaño incrustados indican claramente que esta especie era un animal social que vivía en manadas. Los deinonicos se valían de su gran número para cooperar en la caza y dar muerte así a animales mucho mayores que ellos. Parece que el gran herbívoro Tenontosaurus de hasta 6,5 metros de longitud (más del doble que un deinonico adulto) era una presa habitual en su dieta.
Tras haber rodeado a la víctima con la ayuda de ingeniosas tácticas de caza, los deinonicos saltaban sobre ella y procedían a desgarrarla con sus uñas afiladas, especialmente la gran garra en forma de hoz de cada pie, que era retráctil y podía golpear bastante fuerte gracias a los potentes músculos que la controlaban. Los dientes los usaban en raras ocasiones para matar a la víctima; los que se han encontrado incrustados sobre los huesos de tenontosaurios seguramente acabaron ahí al devorar a la presa. Se ignora qué clase de organización habría en las manadas de deinonico, pero al ser depredadores sociales capaces de tramar estrategias de caza, es muy probable que tuviesen una organización jerárquica similar a la de los cánidos o hienas actuales.
[editar] Historia
Antigua visión de Deinonychus sin plumas.Los restos fósiles de Deinonychus han sido recobrados de la Formación Cloverly de Montana y Wyoming[4] y de la Formación Antlers de Oklahoma,[5] en Estados Unidos. Adicionalmente, un diente de las capas de arcilla de Cloveryy de la Formación Potomac en la costa atlántica de Maryland ha sido asignado a este género.[6]
Los primeros restos en ser encontrados, se hallaron en 1931 en el sudeste de Montana cerca del pueblo de Bridger. Un grupo liderado por el , paleontólogo Barnum Brown, se concentraron sobre todo en excavar y preparar los restos del ornitópodo Tenontosaurus, pero en su informe de campo sobre el sitio excavación para el Museo Americano de Historia Natural, informo el descubrimiento de un pequeño dinosaurio carnívoro cerca del esqueleto de Tenontosaurus, "pero muy encajado en la roca lo que lo hacia difícil de preparar".[7] Llamó informalmente al animal "Daptosaurus" e hizo preparaciones para describirlo y lo colocó en la exhibición, pero nunca termino este trabajo.[8] Posteriormente B. Brown volvió de la Formación Cloverly con el esqueleto de un terópodo más pequeño con aparentemente los dientes de gran tamaño, al que nombró informalmente como "Megadontosaurus". John Ostrom, reviso el material varias décadas más tarde, identificando a los dientes como pertenecientes a un Deinonychus, el esqueleto proveniente de un animal totalmente distinto al que nombro Microvenator.[8] Un poco más de treinta años más tarde, en agosto de 1964, el paleontólogo John Ostrom llevó a una expedición del Museo Peabody de la Universidad de Yale que descubrió más material esquelético. Las expediciones durante los dos veranos siguientes desenterraron más de 1000 huesos, entre los cuales estaban por lo menos tres individuos. Puesto que la asociación entre los varios huesos recuperados era débil, la fabricación del número exacto de animales individuales se presentó imposible de determinar correctamente, el espécimen tipo (YPM 5205) de Deinonychus fue restringido a un pie izquierdo completo y un pie derecho parcial que pertenecieron definitivamente al mismo individuo.[9] Los especímenes restantes fueron catalogados en cincuenta entradas separadas en el Museo Peabody de Historia Natural deYale.
Posteriores estudios de J. Ostrom y G. E. Meyer analizaron su propio material así como el "Daptosaurus" de B. Brown detalladamente y encontraron que era lo misma especie. J. Ostrom publico su descubrimiento en 1969, dandole a todos los restos referido al nombre de Deinonychus antirrhopus.[9]
Aunque una miríada de huesos estaba disponible antes de 1969, muchos importantes faltaban o eran difíciles de interpretar. Había pocos elementos postorbitales del cráneo, ningún fémures, sacro, espoleta o esternón, vértebras faltantes, y J. Ostrom pensó que solamente un fragmento minúsculo era el coracoides. La reconstrucción esquelética de J. Ostrom de Deinonychus incluyó un hueso pélvico muy inusual, un pubis trapezoidal y plano, diferente al de los otros terópodos, pero que era de la misma longitud que el isquion y que fue encontrada a la derecha de este.
[editar] Posteriores descubrimientos
En 1974 J. Ostrom publicó otra monografía sobre el hombro de Deinonychus en cuál corrigió que el pubis que él había descrito era realmente un coracoides, un elemento del hombro.[10] En ese mismo año, otro espécimen de Deinonychus fue excavado en Montana por una expedición de la Universidad de Harvard dirigida por Farish Jenkins. Este descubrimiento agregó varios nuevos elementos; fémures bien preservados, pubis, un sacro, y mejores ilion, así como los elementos del pie y del metatarso.J. Ostrom describió este espécimen y revisó la restauración esquelética de Deinonychus. Esta vez se vieron pubis muy largos, y J. Ostrom comenzó a sospechar que pudieron incluso haber estado orientados hacia cauda poco como en los pájaros.[11]
Representación del Deinonychus con plumas.Un esqueleto de Deinonychus que incluye los huesos del especimen original y más completo puede verse en el Museo Americano de Historia Natural,[12] con otro ejemplar en el Museo de Zoología Comparada de la Universidad de Harvard. Los especímenes del Museo Americanos y de Harvard son de un lugar distinto que los de Yale. Incluso estos dos montajes esqueléticos están carecen de elementos incluyendo los esternones, las costillas esternales, la espoleta, y gastralias.
Incluso después de todo el trabajo de J. Ostrom, varios pequeños bloques de material incrustado en la roca seguía estando sin preparación en el almacén del AMNH. Éstos consistieron sobre todo en huesos y fragmentos aislados de huesos, incluidos en la matriz original, o en la roca circundante en la cual los especímenes terminaron enterrados inicialmente. Un examen de estos bloques sin preparación de Gerald Grellet-Tinner y de Peter Makovicky en 2000 reveló una característica interesante, pasada por alto. Varios huesos largos y finos identificados en los bloques como tendones osificados (estructuras que ayudaron a atiesar la cola de Deinonychus) resultado representar realmente gastralias (costillas abdominales). Más perceptiblemente, una gran cantidad de cáscaras de huevo fosilizadas previamente inadvertidas fueron descubiertas en la matriz de la roca que había rodeado el espécimen original de Deinonychus.[13]
En un informe posterior, más detallado sobre las cáscaras de huevo, G. Grellet-Tinner y P. Makovicky concluyeron que los huevos pertenecieron casi ciertamente a Deinonychus, representando el primer huevo del dromeosáurido que se identificará.[7] Por otra parte, la superficie externa de una cáscara de huevo fue encontrada en contacto cercano con las gastralias que sugerriria que ese Deinonychus pudo haber estado incubando sus huevos. Esto implica que Deinonychus realizaba un traspaso térmico del cuerpo como mecanismo para la incubación del huevo, e indica endotermia similar a los pájaros modernos.[14] El estudio adicional de Gregory Erickson y colegas encuentra que este individuo tenia entre 13 o 14 años a la muerte y su crecimiento amesetado. A diferencia de otros de otros terópodos en su estudio de especímenes encontrados asociado con los huevos o nidos, había acabado el crecimiento a la hora de su muerte. [15]
[editar] Implicaciones
La descripción de 1969 hecha por J. Ostrom de Deinonychus ha sido descrito como uno de los hechos más importantes de la paleontología del mediados del siglo XX.[16] El hallazgo de los primeros restos de muchos de Deinonychus por los paleontólogos John Ostrom y Grant E. Meyer en 1964, en el sur de Montana, llevaron a Ostrom a suponer que el Deinonychus vivía en manadas, algo impensable para la época, que juzgaba a los dinosaurios tan obligádamente estúpidos que sólo podrían llevar una vida solitaria. También dio con las primeras evidencias de que los deinonico depredaban sobre los Tenontosaurus, algo que, evidentemente, no podrían hacer en solitario.
Esqueleto de un deinonico.Uniendo estas evidencias con su singular anatomía, Ostrom describió al Deinonychus como un animal activo, ágil, veloz y mínimamente inteligente, características que implicaban necesariamente el que fuera un animal de sangre caliente. Tal idea tuvo un fuerte impacto en los círculos académicos y posteriormente en la idea popular sobre los dinosaurios, que llevaron a lo que se ha denominado como Renacimiento de los Dinosaurios, el abandono de los prejuicios decimonónicos sobre los dinosaurios y su estudio sistemático como animales de su tiempo. La avalancha de descubrimientos posteriores que ha dibujado a los dinosaurios como animales bien adaptados, inteligentes, ágiles, de sangre caliente, buenos padres de su prole y capaces de tener vida social, serían impensables si el Deinonychus no hubiese llevado a Ostrom a dar el pistoletazo de salida. Varios años después, J. Ostrom noto las similitudes entre los miembros de Deinonychus con las aves, siendo esta observación lo que le llevo a revivir la hipótesis que las aves descendían de los dinosaurios.[17] Treinta años después esa idea esta mundialmente aceptada.
Deinonychus estaba seguramente emplumado como su cercano pariente Velociraptor, demostrado en 2007 en base de las marcas de las plumas en los brazos. Tales marcas demuestran donde se anclan las plumas en pájaros modernos. Este descubrimiento reciente agregó el peso a las teorías ya fuertes que todos los dromeosáuridos fueron emplumados.[18]
[editar] Clasificación
Varios años después, el estudio de los fósiles de Deinonychus permitió a Ostrom apreciar ciertas similitudes entre éste y las aves. Con esto, Ostrom propuso que las aves descendían de los dinosaurios, algo que ya había mencionado Thomas Henry Huxley en el siglo XIX pero que había caído en el olvido desde los años 20 del siglo pasado. Pese a las fuertes críticas de los ornitólogos y paleontólogos de la época, decenas de descubrimientos en los 30 años siguientes han dado la razón a Ostrom. Hoy en día, incluso se conocen numerosos deinonicosaurios y dinosaurios similares con plumas avianas auténticas como Velociraptor o Microraptor, por lo que es muy probable que el propio Deinonychus tuviera también una cubierta plumosa que le ayudase a regular el calor corporal. Deinonicosaurios (Deinonychosauria) y aves (Avialae) están reunidas actualmente en el clado Maniraptora.
Esqueleto de un D. antirrhopus.Deinonychus es uno de los mejor conocidos entre los Dromaeosauridae,[19] junto al más pequeño y relativamente cercano Velociraptor, del Campaniano de Asia Central.[20] [21] El clado que ellos forman se llama Velociraptorinae. El nombre Velociraptorinae fue por primera vez acuñado por Rinchen Barsbold en 1983[22] y originalmente contenía solo al Velociraptor. Posteriormante Phil Currie incluyo a más dromeosáuridos.[23] Del Cretácico superior a Tsaagan de Mongolia[20] y al norteaméricano Saurornitholestes,[2] pueden también estar entre los familiares cercanos, pero este último es mal conocido y difícil de clasificar.[20] Velociraptor y miran a sus parientes usaron sus garras más que sus cráneos como herramientas de caza, en comparación con dromeosáuridos como Dromaeosaurus de cráneos mas masivos.[24] Junto con Troodontidae, los dromeosáuridos del clado Deinonychosauria son taxones hermanos de las aves. Filogenícamente, Deinonychosauria al grupo de dinosaurios no aviares más cercano a las aves.[25]
[editar] Paleoecología
La evidencia geologica sugiere que Deinonychus habitaba una llanura aluvional o zonas pantanosas.[19] El paleoambiente de ambas formaciones, la Cloverly y la de Antlers, en dondes los restos de Deinonychus han sido encontrados, consisten de bosques, deltas fluviales y lagos, no muy diferente a lo que es hoy Louisiana.[26] Otros animales que compartieron este mundo con Deinonychus al herbívoro armado Sauropelta y los ornitópodos Zephyrosaurus y Tenontosaurus. En Oklahoma, el ecosistema de Deinonychus también incluyo al terópodo Acrocanthosaurus, y el gigante Sauroposeidon, el cocodriliano Goniopholis, y el Lepisosteiformes Lepisosteus.[26] Si los dientes encontrados en Maryland son de ' ' Deinonychus' ' , entonces sus vecinos incluirían Astrodon , Dryptosaurus y un nodosáurido, llamado Priconodon conocido solamente por los dientes.
[editar] Paleobiología
[editar] Comportamiento predador
cabeza de Deinonychus basado en la reciente reconstrucción del cráneo.Los dientes de Deinonychus encontrados en asociación con fósiles del dinosaurio ornitópodo Tenontosaurus son muy comunes en la Formación Cloverly. Se han descubierto dos sitios que preservan fósiles bastantes completos de Deinonychus cerca a Tenontosaurus. El primero, el sitio de Yale en Cloverly, Montana, incluye numerosos dientes, cuatro fósiles de Deinonychus adultos y un juvenil. La asociación de este número de esqueletos de Deinonychus en una solo lugar sugiere que Deinonychus puede haberse alimentado de ese animal, y quizás haberlo cazado. J. Ostrom y W. Maxwell incluso han utilizado esta información para especular que Deinonychus pudo haber vivido y cazado en manadas.[27] El segundo Sitio es en la Formación Antlers de Oklahoma. El sitio contiene seis esqueletos parciales de Tenontosaurus de varios tamaños, junto con un esquelético parcial y muchos dientes de Deinonychus. Un húmero del Tenontosaurus incluso tiene lo qué pudo ser marcas de dientes de Deinonychus. D. Brinkman et al. . (1998) precisan que Deinonychus tenía un peso adulto de 70 a 100 kilogramos, mientras que los Tenontosaurus adultos pesaban entre 1 a 4 toneladas. Un solitario Deinonychus no podría matar un Tenontosaurus adulto, sugiriendo que la caza en manadas sería posible.[28]
Un estudio reciente por B. T. Roach y D. L. Brinkman han aportado en la pregunta sobre el comportamiento cooperativo durante la caza en manada de Deinonychus , basados en lo que se sabe de la caza de carnívoros modernos y tafonomía de sitios de Tenontosaurus. Los arcosaurios modernos (pájaros y cocodrilos) y el Dragón de Komodo exhiben poco de caza cooperativa, en su lugar, son generalmente cazadores solitarios, o se agrupan en carcasas muertas anteriormente, donde ocurre mucho conflicto entre los individuos de la misma especie. Por ejemplo, en las situaciones donde los grupos de dragones de Komodo están comiendo juntos, los individuos más grandes comen primero y atacarán a Dragones de Komodo más pequeños que intenten alimentarse. Si se mata al animal más pequeño, éste es canibalizado. Cuando esta información se aplica a los sitios de Tenontosaurus, parece que es consistente con lo encontrado entre los Dragones de Komodo y cocodrilos. Los restos esqueléticos de Deinonychus encontrados en estos sitios son de subadultos y las piezas que faltan en ellos son consistentes con la predación por otro Deinonychus.[29]
[editar] Función de los miembros
A pesar de ser la característica más distintiva de Deinonychus, la forma y la curvatura en hoz de la garra varía entre los especímenes. El espécimen tipo descrito por J. Ostrom en 1969 tiene una garra fuertemente curvada en hoz, mientras que un espécimen más nuevo descrito en 1976 tenía una garra con una curvatura mucho más débil, más similar en perfil con el de las garras normales en los restantes dedos del pie.[30] J. Ostrom ha sugerido que esta diferencia en el tamaño y la forma de las garras en hoz podría ser debido a la variación individual, sexual, o relativa a la edad.
Modelo del los huesos del Deinonychus mostrando la garra falciforme, en el Museo de Zoología de Copenhagen.J. Ostrom especuló originalmente que Deinonychus agarraba sus presas con las garras de las manos mientras que pateaba desgarrando con sus garras falciformes.[9] Posteriores estudios, sin embargo, han demostrado que las garras falciformes no fueron utilizadas para desgarrar pero si para asestar pequeñas puñaladas a la víctima,[31] posiblemente indicando su uso como crampones para escalar.[32] Estudios biomecánicos Ken Carpenter en 2002 confirmaron que la función más probable de los brazos era la de agarrar la presa, pues sus grandes longitudes habrían permitido un alcance más largo que para la mayoría de los otros terópodos. El grande y alargado coracoides hace suponer la existencia de músculos de gran alcance en los brazos, lo que consolida esta teoría.[33] Los estudios biomecánicos de K. Carpenter usando moldes de los huesos también demostraron que los Deinonychus no podían doblar sus brazos contra su cuerpo como un pájaro, contrariamente lo qué fue deducido en 1985 de las descripciones anteriores por Jacques Gauthier[34] y Gregory S. Paul en 1988.[2]
Ilustración de Deinonychus antirrhopus en descanso con los brazos doblados.
Plumaje basado en géneros relacionados.Los estudios de Phil Senter en 2006 indicaron que los brazos de Deinonychus se podían utilizar no sólo para agarrar sino también para llevar los objetos hacia el pecho. La presencia probable de un ala grande emplumada en Deinonychus , sin embargo, habría limitado en un cierto grado la gama de movimiento de los brazos. Por ejemplo, cuando Deinonychus llevaba sus brazos hacia adelante, la palma de la mano giraba automáticamente a una posición hacia arriba. Esto habría hecho que un ala bloqueara a la otra si ambos brazos estuviesen extendidos al mismo tiempo, llevando P. Senter a concluir eso que lo atraer objetos al pecho habría sido lograda solamente con un brazo a la vez. La función de los dedos también habría estado limitada por las plumas; por ejemplo, solamente el tercer dígito de la mano se habría podido emplear en actividades tales escarbar en las grietas para buscar pequeñas presa, y solamente en perpendicular a la posición del ala.[35] Alan Gishlick, en un 2001 estudio de la mecánica del brazo de Deinonychus, encontró que incluso si las plumas grandes del ala estaban presentes, la capacidad de atrape de la mano no habría sido obstaculizada perceptiblemente. Esta habría sido perpendicular al ala, y habría sido sostenida probablemente por ambas manos simultáneamente en un " abrazo de oso" de manera, hecho que ha sido apoyado por los estudios posteriores del brazo por Carpenter y Senter.[36]
W. Parsons ha demostrado que los especímenes juveniles y subadultos de Deinonychus exhiban algunas diferencias morfológicas con los adultos. Por ejemplo, los brazos de los especímenes más jóvenes eran proporcional más largos que los de los adultos, una indicación posible de la diferencia en comportamiento entre los jóvenes y adultos.[37]
[editar] Velocidad
Buitreraptor y Deinonychus montados en posición de correr.Los dromeosáuridos, especialmente Deinonychus, se representan a menudo como animales inusualmente de rápidos en los medios masivos, y J. Ostrom mismo especuló que Deinonychus era veloz en su descripción original.[9] Sin embargo, cuando el primer ' ' Deinonychus' ' fue descrito, carecia de una pierna completa, y J. Ostrom especulo sobre la longitud del fémur, que más adelante se comprobo que resulto una sobrestimación. En un estudio posterior, J. Ostrom observó que cociente del fémur a tibia no es tan importante en la determinación de velocidad como la longitud relativa del pie y de la parte inferior de la pierna. En los pájaros modernos, como el avestruz, el cociente entre el pie y la tibia es 0,95. En dinosaurios inusualmente de rápidos como el Struthiomimus , el cociente es 0,68, pero en Deinonychus , el cociente es de 0,48. J. Ostrom indicó que "solamente una conclusión es razonable, el Deinonychus no era particularmente rápido comparado con otros dinosaurios, ni con las aves corredoras modernas".[30]
El bajo cociente del miembro posterior distal en Deinonychus es debido en parte a un inusuálmente corto metatarso. El cociente es realmente más grande en individuos más pequeños que en los más grandes. J. Ostrom sugirió que el metatarso corto se pueda relacionar con la función de la garra falciforme, y utilizó el hecho de que parece hacerse más corto mientras que los individuos envejecen como soporte de esto, Interpretó todas estas características, el segundo dedo del pie corto con la garra agrandada, el metatarso corto, etc. como ayuda para el uso de la pierna trasera como arma ofensiva, donde la garra giraría hacia abajo y atras, y de la pierna empujaba hacia abajo y detrás al mismo tiempo, desgarrando en la presa. J. Ostrom sugirió que el metatarso corto redujera la tensión total en los huesos de la pierna durante tal ataque, e interpretó el arreglo inusual de las inserciones musculares en la pierna del Deinonychus como apoyo para su idea que un sistema de músculos distinto fuera utilizado en el movimiento rapaz y en caminar. Por lo tanto, J. Ostrom concluyó que las piernas de Deinonychus representaban un equilibrio entre las adaptaciones corrientes necesarias para un depredador ágil, y las características para compensar su arma única del pie.[30]
En su estudio de 1981 de las huellas de dinosaurio encontrada en Canadá, Richard Kool realizo estimaciones groseras sobre la velocidad que caminan basadas en varios rastros de huellas hechas por diversas especie en la Formación Gething de la Columbia Británica. R. Kool estimo para una de estos rastros de icnospecies, Irenichnites gracilis (que pudo haber sido de hecho Deinonychus), una velocidad al caminar de 2,8 metros por segundo (alrededor de 10,1 km por hora).[38]
[editar] Huevos
La identificación en 2000 de un probable huevo de Deinonychus asociado a uno de los especímenes originales permitió la comparación con otros dinosaurios terópodos en términos de estructura del huevo, nido, y reproducción. En su estudio de 2006 del espécimen, G. Grellet-Tinner y P. Makovicky examinaron la posibilidad que los dromeosáuridos se había estado alimentando de los huevos, o que fragmentos había sido asociado al esqueleto de Deinonychus por coincidencia. Descartaron la idea que el huevo había sido una comida para el terópodo, observando que los fragmentos fueron intercalados entre las gastralias y los huesos del brazo, haciéndolo imposible que representaran el contenido estomacal del animal. Además, la manera de la cual el huevo había sido machacado y hecho fragmento indicando que había estado intacto a la hora del entierro, y estaba quebrado por el proceso de fosilización. La idea que el huevo fuera asociado aleatoriamente al dinosaurio también fue encontrada como inverosímil, los huesos que rodeaban el huevo no habían sido dispersados ni desarticulado, sino seguían estando bastante intactos concerniente a sus posiciones en vida, indicando que el área alrededor al huevo no fueron disturbadas durante la preservación. El hecho de que estos huesos fueran gastralias, que muy raramente se encuentran articulados, apoyó esta interpretación. Toda la evidencia, según G. Grellet-Tinner y P. Makovicky, indican que el huevo estaba intacto debajo del cuerpo del Deinonychus cuando fue enterrado. Es posible que éste representa el empollamiento o comportamiento de nido en Deinonychus similar a lo visto en troodóntidos relacionados y oviraptóridos, o que el huevo estaba de hecho en el interior del oviducto cuando murió el animal.[7]
Deinonychus echado en el suelo.El examen de la microestructura del huevo de Deinonychus confirma que perteneció a un terópodo, puesto que comparte características con otros huevos conocidos de terópodos y muestra que diferencias con huevos de ornitisquios y saurópodos. Comparado a otro terópodos maniraptores, el huevo de Deinonychus es más similar a los de oviraptórido que a los de troodóntidos, a pesar de los estudios que demuestran que estos últimos son lo más estrechamente vinculados a los dromeosáuridos como ' ' Deinonychus' '. Mientras que el huevo estaba muy roto para determinar exactamente su tamaño, G. Grellet-Tinner y P. Makovicky estimaban un diámetro alrededor de 7 cm basado en la anchura del canal pélvico a través del cual el huevo tuvo que haber pasado. Este tamaño es similar al diámetro de 7.2 cm del más grande huevo de Citipati huevos de un oviraptórido. Citipati y Deinonychus también compartieron el mismo tamaño de cuerpo total, apoyando esta estimación. Además, el grosor de las cáscaras de huevo de Citipati y Deinonychus son casi idénticas, y puesto que el grueso de cáscara se correlaciona con el volumen del huevo, estos datos apoyan la idea que los huevos de estos dos animales eran de tamaño casi igual.[7]
El Stegosaurus más grande conocido llego a medir 12 metros de largo, 4 metros de altura y un peso de 3 toneladas. El tamaño promedio de Stegosaurus es de alrededor 9 metros de largo y de 4 metros de alto, el Stegosaurus es uno lo más fácilmente identificables de los dinosaurios cuadrúpedos, debido a la doble fila distintiva de las placas romboidales que se elevan verticalmente a lo largo de su arqueado lomo y de los dos pares de púas largas que se extienden horizontalmente cerca del extremo de su cola. Aunque fue un animal grande, quedaba empequeñecido por sus contemporáneos los saurópodos gigantes como Diplodocus, Camarasaurus y Apatosaurus. Una cierta forma de armadura aparece haber sido necesaria ya coexistió con los dinosaurios de terópodos cazadores grandes, tales como los temibles Allosaurus y el Ceratosaurus.
Modelo de Stegosaurus en el Parque del Jurásico en Baltów, Polonia.El cuerpo, sostenido de forma obligada por cuatro patas acabadas en pezuñas, se estrechaba y achicaba hacia delante, hasta terminar en una minúscula cabeza alargada dotada de un pico débil y dientes pequeños de escasa capacidad masticatoria. Los pies traseros tenían tres dedos cortos en cada pie, mientras que cada pata delantera tenía cinco dedos del pie, solamente los dos dedos del pie internos tenían una forma de casco. Los cuatro miembros se apoyaban en almohadillas detrás de los dedos del pie.[3] Los miembros delanteros eran mucho más corto que los rechonchos miembros traseros, que dieron lugar a una postura inusual. La cola aparece haber sido sostenida bien lejos de la tierra, mientras que la cabeza de Stegosaurus tenia una posición cerca del suelo, probablemente no más alto que 1 metro sobre la tierra. [4]
Esqueleto de Segosaurus.El largo y angosto cráneo era pequeño en proporción del cuerpo. Tenia una pequemña fenestra antorbital, el agujero entre la nariz y el ojo común a la mayoría arcosaurios, incluyendo los pájaros modernos, aunque perdido en cocodrilos actuales. El cráneo en la posición baja sugiere que Stegosaurus puede haber sido un ramoreador de la vegetación de crecimiento bajo. Esta interpretación es apoyada por la ausencia de dientes delanteros y su reemplazo por un pico córneo o rhamphotheca. Los dientes de Stegosaurus eran pequeños, triangulares y con las facetas planas de desgaste demuestran que molieron su alimento. La colocación de la inserción en las quijadas sugiere que Stegosaurus tenía mejillas para mantener el alimento sus bocas mientras masticaban.[5]
A pesar del tamaño total del animal, la caja craneana de Stegosaurus era pequeña, siendo no más grande que la de un perro. Una bien conservada caja craneana deStegosaurus permitió a Othniel Charles Marsh obtener un molde en 1880 de la cavidad del cerebro del animal, que dio una indicación del tamaño del cerebro. El este molde demostró que el cerebro era de hecho muy pequeño, quizá el más pequeño entre los dinosaurios. El hecho de que un animal que pesaba sobre 4,5 toneladas podría tener un cerebro no más de 80 gramos de contribuidos a la vieja idea popular que los dinosaurios eran estúpidos, una idea ahora rechazada en gran parte.[6] Ya en el siglo XIX se convirtió en el paradigma del dinosaurio estúpido, debido a que su cerebro era poco más grande que una nuez. Aunque hoy se sabe que el cerebro no crece de forma proporcional al resto del cuerpo y que en la mayoría de los dinosaurios tenía un tamaño normal para éstos, los estegosaurios lo siguen teniendo ligeramente más pequeño de lo que sería esperable. Así pues, no debían tener los sentidos muy desarrollados.
Comparativa de tamaño entre un estegosaurio y un hombre.Las características más notables de este animal son sus cuatro púas de hasta 60 centímetros en la cola y sobre todo, la hilera de anchas placas que coronaban su espalda. Su posición ha sido muy discutida en tiempos pasados, llegando a sugerirse que se distribuían paralelas sobre el lomo o incluso pegadas al cuerpo a modo de armadura. No obstante, los conocimientos actuales permiten asegurar que se distribuían en dos filas sobre el lomo, de forma alterna y en posición vertical. Su papel defensorio era despreciable debido a su escaso grosor y fuerte vascularización, así que probablemente tenían un papel más disuasorio que otra cosa. Un estudio de McWhinney et al. en el 2001,[7] dio a conocer un alto grado de traumas en las púas de la cola, sugiriendo que estas eran usadas como defensa. Erguidas sobre el lomo del animal, éste parecería más grande aún a los depredadores como el alosaurio y el ceratosaurio. Se ha sugerido recientemente que la vascularización de las mismas podría teñir las placas de vivos colores a voluntad del animal, bombeando sangre con fuerza hacia ellas, y pudiendo así tener un papel en el cortejo o aumentando su poder disuasorio.
Otra teoría las considera un instrumento termorregulador. Los dinosaurios son considerados animales de sangre caliente hoy en día, así que las placas no servirían para calentar el animal como se creyó en su momento, sino para enfriarlo. Dado que no masticaba su alimento, el estegosaurio tragaba enormes cantidades de plantas que fermentaban dentro de sus grandes intestinos: esto originaba enormes cantidades de calor que eran emitidas al exterior por las placas, salvando al animal de achicharrarse con su propio metabolismo. Un experimento con una maqueta que reproducía la forma de un Stegosaurus corroboró esta idea. También se observa que las placas son progresivamente más anchas según asciende el tamaño entre los miembros de la familia Stegosauridae, de la que esta especie es su mayor representante.
La mayor parte de la información conocida sobre Stegosaurus viene de restos de animales maduros, sin embargo restos más recientemente de juveniles de Stegosaurus se han encontrado. Un espécimen subadulto, descubierto en 1994 en Wyoming, mide 4,6 metros de largo y 2 metros de alto, y se estima para haber pesado 2,3 toneladas en vida. Está en la exhibición en el museo geológico de la universidad de Wyoming.[8] El esqueleto más pequeño conocido, mide 210 centímetros de largo y 80¢ímetros de alto en el lomo, se exhibe en el Muceoi de Naturaleza y Ciencia de Denver.
[editar] Historia
Especimen fósil de S. stenops mostrado como fue encontrado.Stegosaurus, uno de los muchos dinosaurios recogidos y descritos por primera vez en la llamada Guerras de los huesos, fue nombrado originalmente por Othniel Charles Marsh en 1877,[9] a partir de restos encontrados al norte de Morrison, Colorado. Estos primeros restos son el holotipo de Stegosaurus armatus. La bases del nombre científico, lagarto con tejado se ha pensado fue colocado por Marsh en la creencia inicial de que las placas cubrían completamente el lomo del animal, traslapándose como teja s en un techo. Una gran cantidad de material de Stegosaurus fue recuperado durante los siguientes años y Marsh publicó varios trabajos sobre el género. Inicialmente, varias especies fueron descritas. Sin embargo, muchos de éstas se han considerado desde entonces como inválidas o sinónimos de la especie existente, [10] dejando solo dos especies bien conocidas y una pobremente. Los restos de Stegosaurus se han encontrado en la Formación Morrison , en las las zonas estratigráficas 2 a 6, con material adicional posiblemente referible a Stegosaurus recuperado de la zona estratigráfica 1.[11]
[editar] Especies válidas
Locaciones
S. armatus
Formación Morrison, Colorado, Wyoming, y Utah, EE.UU.[12]
S. stenops
Formación Morrison, Colorado, Wyoming, y Utah, EE.UU.[12]
S. longispinus
Formación Morrison, Wyoming y posiblemente Utah, EE.UU.[12]
Stegosaurus armatus: significa "lagarto con tejado armado", es la primera especie que se encontrará y se conoce a partir de dos esqueletos parciales, de dos cráneos parciales y por lo menos de treinta individuos fragmentarios.[9] Esta especie tenía cuatro púas horizontales en la cola y placas relativamente pequeñas. Media alrededor de 9 metros siendo la mayor de las especies de Stegosaurus.
Dibujo en lápiz de un Stegosaurus de 1923.Stegosaurus stenops: significa "lagarto con tejado estrecho", fue nombrado por Marsh en 1887,[13] a partir del holotipo recolectado por by Marshal Felch enGarden Park, al norte de Cañón City, Colorado, en 1886. Esta es la mejor conocida de las especies de Stegosaurus, principalmente porque sus restos incluyen por lo menos un esqueleto articulado completo. Tenía las placas grandes, amplias y cuatro púas en la cola. Stegosaurus stenops es conocido de por lo menos de 50 esqueletos parciales de adultos y jóvenes, un cráneo completo y cuatro cráneos parciales. Era más corto que S. armatus, midiendo 7 metros.
Stegosaurus longispinus: significa "lagarto con tejado de espinas largas", fue nombrado por Charles W. Gilmore[14] y conocido por un esqueleto parcial, de la Formación Morrison en Wyoming. Stegosaurus longispinus era notable para su sistema de cuatro espinas de la cola inusualmente largas. Algunos la consideran una especie de Kentrosaurus. Como S. stenops, midió unos 7 metros.
[editar] Especies dudosas
Stegosaurus ungulatus: significa "lagarto con tejado con cascos", fue nombrado por Marsh en 1879, a partir de restos recobrados en Como Bluff, Wyoming.[15] It is known from a few vertebrae and armor plates. It might be a juvenile form of S. armatus,[16] aunque el material original de S. armatus todavía espera ser descrita completamente. El espécimen descubierto en Portugal y datado de la parte superior Kimmeridgiano o inferior del Titoniano se ha atribuido a esta especie.[2]
Stegosaurus sulcatus: significa "lagarto con tejado con surcos" fue descrito por Marsh en 1887 basado en un esqueleto parcial.[13] Es considerado sinónimo de S. armatus.[16]
Stegosaurus, dibujo de 1902.Stegosaurus duplex: significa "lagarto con tejado con dos plexos", en la alusión al canal neural agrandado en el sacro que MArsh presento como un caso de "cerebro posterior del ", es probablemente sinónimo de S. armatus.[16] Aunque fueron nombrados por Marsh en 1887, incluyendo los especímenes holotipo, los huesos desarticulados fueron recogidos realmente en 1879 por Edward Ashley en Como Bluff, Wyoming.
Stegosaurus seeleyanus, originalmente llamados Hypsirophus, es probablemente el mismo que S. armatus.
Stegosaurus (Diracodon) laticeps fue descrito por Marsh en 1881, a partir de algunos fragmentos de la mandíbula inferiro.[17] Esto hace que algunos consideren a S. stenops una especie de Diracodon, y otros a Diracodon en si como especie de Stegosaurus. Bakker ha retomado a D. laticeps en 1986,[18] aunque otros observen que el material es no diagnóstico y probablemente sinónimo con S. stenops.[10]
Stegosaurus affinis, descrito por Marsh in 1881, es solo conocido por un pubis y considerado como dudoso.[16] Es posiblemente sinónimo de S. armatus.[14]
[editar] Especies reasignadas
Stegosaurus madagascariensis de Madagascar es solo conocida por un diente y descrita por Piveteau en 1926. El diente ha sido alternativamente asignado a un estegosáurido, al terópodo Majungasaurus,[19] a un hadrosáurido o hasta un cocodrilo.
Stegosaurus, dibujo antigu

tros restos una vez asignados a Stegosaurus hoy son reconocidos como géneros diferentes. Este es el caso de Stegosaurus marshi, descrito por Lucas en 1901. Fue renombrado Hoplitosaurus en 1902. Stegosaurus priscus, descrito por Nopcsa en 1911, y posteriormente reasignado a Lexovisaurus,[16] y hoy considerado una especie de Loricatosaurus.[20]
[editar] Clasificación
Stegosaurus fue el primer género nombrado de la familia Stegosauridae. Es el género tipo que le dio el nombre a esta familia. Stegosauridae es una de las familias dentro del infraorden Stegosauria, junto con Huayangosauridae. Stegosauria pertenece a Thyreophora, un grupo de dinosaurios armados, un suborden que comparte con los más variados Ankylosauria. Los estegosáuridos on un clado de animales de apariencia similar en postura y forma de corporal, diferenciándose principalmente en el arreglo de placas y espinas del cuerpo. Los parientes más cercanos a Stegosaurus son Wuerhosaurus de China y Kentrosaurus del este de África.
[editar] Orígines
Los orígenes de Stegosaurus son inciertos, con pocos restos de estegosáuridos basales y de sus antepasados conocidos. Recientemente, los estegosáuridos se han demostrado como presentes en la parte más baja de la Formación Morrison, existiendo varios millones de años antes de la aparición de de Stegosaurus en si, con el descubrimiento del cercano Hesperosaurus de principios del Kimeridgiano.[21] El más antiguoi estegosáurido conocido Lexovisaurus, proviene de la Formación Arcilla de Oxford de Inglaterra y Francia, lo que le da una edad de principios o mediados del Calloviano.
S. ungulatus en el Museo Carnegie de Historia Natural.Los géneros más basales y tempranos como Huayangosaurus del Jurásico Medio en China, alrededor de 165 millones de años, precede a Stegosaurus por 20 millones de años y es un género de la familia Huayangosauridae. Aún más temprano es Scelidosaurus, del Jurásico Inferior de Inglaterra, que vivió hace aproximadamente 190 millones de años. Es interesante que presente características de los anquilosaurianos y los estegosaurianos. Emausaurus de Alemania era otro pequeño cuadrúpedo, mientras que Scutellosaurus de Arizona en los E.E.U.U. era incluso un género anterior y facultativo bípedo. Estos dinosaurios pequeños, ligeramente acorazados estaban estrechamente vinculados al antepasado directo de los estegosaurianos y de anquilosaurianos. Un rastos de huellas posiblemente pertenecientes a un dinosaurio acorazado temprano, datada hace 195 millones de años, se ha encontrado en Francia.[22]
[editar] Paleobiología
Stegosaurus junto a la vegetación típica del jurásico.Stegosaurus es el mayor de los estegosáuridos, alcanzando los 12 metros de largo y posiblemente unos 5 000 kilogramos de peso. Sin embargo, 7 a 9 era el largo usual para estos animales. Apenas después del descubrimiento, Marsh considero a Stegosaurus como un [[bípedo, debido a sus miembros delateros relativamente cortos.[23] Él mismo cambio de idea por 1891, para considerarlo un robusto animal cuadrúpedo pesadamente construido.[24] Aunque Stegosaurus ahora se considera indudablemente como un cuadrúpedo, ha habido una cierta discusión si se habría podido alzarse sobre sus piernas traseras, usando su cola para formar un trípode con sus miembros traseros y comer del follaje más alto.[16] esto fue propuesto por Bakker[18] [25] y refutado por Carpenter.[4]
Stegosaurus tenia miembros delanteros muy cortos, con respecto a sus piernas traseras. Además, en sus miembros traseros, la sección inferior compuesta por la tibia y el peroné era muy corta comparada con el fémur. Esto sugiere que no podía correr muy rápido, con la mayor zancada de las de las piernas traseras se habrían mezclado con las piernas delanteras, dando una velocidad máxima de 1,7 a 2 metros por hora (6 a 7 km/h).[5] Rastros de huellas fósiles descubiertos por Matthew Mossbrucker del Museo de Historia Natural de Morrison en Colorado sugiere que los estegosaurios vivían en manadas de animales de todas las edades. Un grupo de huellas se interpreta como cuatro o cinco crías de estegosaurios que se mueven en la misma dirección, mientras que otro tiene una huela de un juvenil estegosaurio con una sobreimpresión de un adulto.[26]
[editar] "Segundo cerebro"
Apenas descripto Stegosaurus, Marsh observó un canal neural agrandado en la región de la cadera, que habría podido acomodar una estructura hasta 20 veces más grande que el cerebro. Esto ha llevado a la idea famosa los dinosaurios como Stegosaurus tenía un “segundo cerebro” en la cola, que pudo haber sido responsable de controlar reflejos en la porción posterior del cuerpo. También se ha sugerido que este “cerebro” pudo haber dado un Stegosaurus una rápida respuesta cuando estaba bajo amenaza de depredadores.[5] Más recientemente, se ha argumentado que este espacio, también encontrado adentro saurópodo s puede haber sido la localización de un cuerpo del glicógeno, una estructura vista en los pájaros actuales de cuya función no se conoce definitivamente pero de los cuales se postula que es para facilitar la fuente glicógeno al sistema nervioso del animal.[27]
[editar] Placas
S. stenops adulto y juvenil, montados bajo el ataque de un Allosaurus, Museo de Ciencia y Naturaleza de Denver.La característica mas reconocible de los estegosaurios son las placas dermales, estas consisten en 17 placas separadas. Estas son osteodermos altamente modificados, similares a los que se ven hoy en día en lagartos y cocodrilos. Estas no estaban directamente conectadas al esqueleto del animal, en su lugar se unian fuertemente a la piel. En el pasado paleontólogos, en especial Robert Bakker, han especulado las placas pudieron haber sido móviles en un cierto grado, aunque otros discrepen con esto.[28] Bakker ha sugerido que las placas fueran los corazones óseos de las agudas placas de cuerno cubiertas que un Stegosaurus podría mover de un tirón de un lado a otro para presentar un depredador un arsenal de los espinas y las láminas afiladas que lo impedirían que se acerque lo suficiente para atacar. Las placas se ubicarían naturalmente a los lados del estegosaurio, la amplitud de las placas que reflejan la anchura del animal en ese punto a lo largo de su espina dorsal. Su razonamiento para que estas placas estén cubiertas de cuerno se basa en que las placas fosilizadas superficialmente tienen una semejanza a los centros óseos de los cuernos en otros animales conocidos o que posiblemente tuvieran cuernos, y para que las placas sean de naturaleza defensivas es que estas tenían una escasa anchura para que se sostenerce erguidas de tal manera que sean útiles en la exhibición sin un esfuerzo muscular continuo.[29] Las placas más grandes fueron encontradas sobre las caderas del animal y medían 60 centímetros de ancho y 60 centímetros de alto. La disposición de las placas ha sido un tema de largo debate, pero la mayoría de los palaeontólogos ahora conviene en que formaron un par de filas alternadas, a cada lado de la línea central de la espalda, lomo, caderas y cola.
Temprana reconstrucción de Stegosaurus con las placas sobre toda la espalda y espinas distribuidas por todo el cuerpo.La función de las placas se ha discutido mucho. Se ensó inicialmente como cierta forma de armadura del lomo del animal.,[23] pero parecen haber sido demasiado frágiles para ser de propósitos defensivos, dejando el anima desprotegido a los flancos.[30] Más recientemente, investigadores han propuesto que contribuían con el control de la temeperatura dela nimal,[28] de una manera similar a la vela del terópodo Spinosaurus o del pelicosaurio Dimetrodon y a las oreyas de los modernos elefantes y liebres. Las placas tenían vasos sanguíneos que corrian a través de surcos y el aire que fluye alrededor de las placas habría refrescado la sangre.[31] Esta teroria ha sido seriamente cuestionada,[32] debido a que los parientes cercanos no tenían esta adaptación y Stegosaurus stenops, tenia una baja superficie de placas y púas implicando que el enfriamiento no era bastante importante para requerir especializaciones de formaciones estructurales tales como placas..
Su gran tamaño sugiere que las placas pudieron haber servido aumentar la altura evidente del animal, para intimidar a cualquiera de sus enemigos[14] o para impresionar a cualquiera de su misma especie, en alguna forma de exhibición,[30] aunque los especímenes machos y hembras parecieran haberlos tenido. Un estudio publicado en 2005 apoya la idea de su uso en la identificación. Los investigadores creen que ésta puede ser la función de otras características anatómicas únicas, encontradas en varia especies de dinosaurios.[33] Stegosaurus stenops también tenía placas en forma de disco en sus caderas.
Uno de los temas más comunes en los libros y trabajos sobre Stegosaurus es su distribución de las placas.[34] La discusión ha sido importante en la historia de la reconstrucción del dinosaurio. Cuatro arreglos posibles de la placa se han debatido durante los años:
Ilustración de Marsh de 1896 de S. stenops. Se ven 12 placas y 8 espinas; Stegosaurus actualmentye tiene 17 y solo 4 espinas.1.Las placas se colocan paralelas a lo largo del lomo del animal, como una armadura. Ésta era la interpretación inicial de Marsh, que llevó al nombre 'lagarto tejado'. Cuando más y mejores placas fueron encontradas, su forma demostró que se colocaban insertaban por un borde impidiendo completamente esta disposición.
2.En 1891, Marsh publico una imagen más popular de Stegosaurus,[24] con una sola linea de placas. Esto fue tempranamente discutido, al parecer porque estaba mal compredido cómo las placas se encajaban en la piel y se creyo que traslaparían demasiado en este arreglo. Fue restablecida, algo modificada, en los años 80, por el artista Stephen Czerkas,[35] basado en el arreglo de las espinas dorsales de la iguana.
3.Las placas se aparearon en una fila doble a lo largo de la espalda. Éste es probablemente el arreglo más común de los cuadros, especialmente anterior unos al 'llamado renacimiento de los dinosaurios en la década de 1970. El estegosaurio de la película de 1933, King Kong tiene este arreglo. Sin embargo, no se ha encontrado dos placas de tamaño y de la forma idénticos en el mismo animal.
4.Dos filas de placas alternadas. Por el principios de los 60, ésta se había convertido en la idea más probable y aun permanece así, principalmente porque un fósil de Stegosaurus el con las placas todavía articuladas indica este arreglo. Una objeción a ella es que este fenómeno es desconocido entre otros reptiles y es difícil entender cómo tal disparidad podría desarrollarse.
[editar] Espinas de la cola (Thagomizer)
Espinas de la cola montadas de un Stegosaurus.Ha habido mucha discusión sobre si las espinas de la cola fueron utilizados para la exhibición solamente, según lo postulado por Gilmore en 1914[14] o usadas como armas. Robert Bakker observó que la cola era probable mucho más flexible que la de otros dinosaurios, ya que careció los tendones osificados, dando crédito a la idea de la cola como arma. Sin embargo, Carpenter[4] ha observado, el traslapo de las placas con muchas de las vértebras de la cola, por lo que el movimiento sería limitado. Bakker también observó que Stegosaurus habría podido maniobrar su parte posterior fácilmente, manteniendo sus grandes miembros tracerso inmóviles y empujando miembros delanteros musculosos pero cortos, permitiendo que girar sobre un eje hábilmente para enfrentarce al atacante.[18] Mas recientemente un estudio de las espinas de McWhinney et al.,[36] el cuál demostró una alta incidencia del daño relacionado con el trauma, presta más peso a la posición que las espinas fueron utilizados de hecho en combate. La ayuda adicional para esta idea era una vértebra perforada de la cola de Allosaurus en cuál entra una espina de la cola perfectamente.[37]
Stegosaurus stenops tiene cuatro espinas en la cola, cada una de entre 60 90centímetros de largo. Descubrimientos de una armadura articulada de estegosaurios, por lo menos en una especie, muestra que las espinas salían horizontalmente de la cola, no verticalmente como se representa a menudo. Inicialmente, Marsh describió a S. armatus como con 87 espinas en la cola, a diferencia de S. stenops. Sin embargo recientes estudios parece confirmar que solo tenían cuatro.[10]
[editar] Dieta
Diagrama de un diente de StegosaurusLos estegosaurios y sus parientes fueron herbívoros. Sin embargo, adoptaron una estrategia de alimentación diferente de la de los otros dinosaurios ornitisquios herbívoros. Los otros ornitisquios poseían dientes capaces de moler el material vegetal y una estructura de quijada capaz de movimientos planos con excepción de simplemente cortar, es decir podrían masticar las plantas. Esto es diferente en el estegosaurio y sus parientes, que tenía pequeños dientes de facetas horizontales de desgaste asociadas al contacto diente-alimento[38] y una mandíbula que solo permitía movimientos de arriba a abajo.[16]
Los estegosaurianos deben haber sido exitosos, con muchas especies y geográficamente dispersos, a finales del Jurásico.[16] Los paleontólogos creen que habría comido plantas tales como musgos, helechos, colas de caballo, los cycas y piñas o frutas.[39] Habrían tragado gastrolitos para ayudar a la digestión de los alimentos, debido a la carencia de la capacidad de masticar, de manera semejante a los pájaros y los cocodrilos modernos.[40] El comer hierbas bajas, común en losmamífero herbívoros modernos de, no habría sido posible para los estegosaurios, ya que los pastos no aparecieron hasta finales del Cretácico cuando los estegosaurios se ya habían desaparecidos.
Una estrategia de alimentación presunta los considera como ramoreadores bajos, comiendo la fruta de varias plantas sin flores, así como el follaje. Este panorama tiene a los estegosaurio comiendo forraje a no más de un metro sobre la tierra.[41] Por una parte, si los estegosaurios habrían podido levantarse en dos piernas, según lo sugerido por Bakker, habrían podido comer de la vegetación y frutas mucho más arriba, con los adultos pudiendo forrajear hasta los 6 sobre la tierra.[5]
[editar] En la cultura popular
Stegosaurus es uno de los más reconocidos dinosaurios.[42] que ha aparecido en infinidad de juguetes, películas, dibujos animados y historietas, formando parte de la visión popular sobre los dinosaurios. Fue parte del renacimiento de los dinosaurios en la década de 1970, donde cambio la forma de ver a lso dinosaurios.[43] Aparece en el libro de Arthur Conan Doyle El mundo perdido
Stegosaurus ha aparecido mucho en el cine, desde luchando con un ceratosaurio en Journey to the Beginning of Time (1954), contra un tiranosaurio, en Planet of Dinosaurs (1978), Walt Disney's Fantasia (1940) donde se ve por primera vez la cola como arma de defensa, también aparece enLand of the Lost (1992-93). Un bebe dinosaurio llamado Spike es uno de los principales personajes de En Busca del Valle Encantado (1988) y sus secuelas.
En el clásico, King Kong (1933), tes el primer animal que los rescatadores ven.Un estegosaurio aparece en El mundo perdido: Parque Jurásico II y aparece brevemente en Parque Jurásico III. Aunque no se ve en la primer película de la serie, su nombre, mal escrito, eaparece en los embriones robados. Estegosaurio es uno de los dinosaurios que usaron para la creación de Godzilla; junto a tiranosaurio e iguanodónte.
También aparece en los documentales The Animal World (1956) combatiendo con un ceratosaurio al igual que en When Dinosaurs Roamed America (2001) y en Jurassic Fight Club (2008) donde combate con un alosaurio, al igual que en el segundo episodio de "Walking with Dinosaurs" (1999), y el especial La balada del gran Al (2000).
Stegosaurus ha aparecido en muchos dibujos animados para niños, entre ellas Transformers Generation 1, En Busca del Valle Encantado, Dinoplativolos, DinoZaurs y Extreme Dinosaurs
En la historieta The Far Side de Gary Larson generalmente se muestran estegosaurios cuando se habla de dinosaurios. De hecho, el termino "Thagomizer" se origina en una tira cómica de Far Side, cuando un profesor cavernícola le enseña a sus alumnos que las espinas de la cola de este animal se llaman de esa manera en honor al "difunto Thag Simmons" que murió atravesado por ellas.[44] El termino ha sido usado por muchos paleontólogos , inclusive en el Instituto Smithsoniano.[45]
El Lambeosaurus mejor conocido, L. lambei , era muy similar al más famoso Corythosaurus en todo menos en la forma del adorno principal de la cabeza. Comparado a Corythosaurus , la cresta de Lambeosaurus estaba más hacia adelate formada por una porción cuadrada mayor apuntando hacia adelante, y una pequeña espina hacia atrás., y los pasos nasales huecos dentro estaban en el frente de la cresta y apilados verticalmente.[1] Se supone que los diferentes tamaños y formas de este órgano dependían de la edad y el sexo (con los más jóvenes careciendo de él). Su cavidad nasal se extendía por dentro de esta cresta mayormente hueca. Se cree que esto pudo haber mejorado el sentido del olfato, o creado un fuerte balido. También puede ser distinguido de Corythosaurus por la falta del proceso nasal bifurcado formando parte de los lados de la cresta, que es la única manera de diferenciar a los jóvenes de los dos géneros, las crestas adquirieron sus formas distintivas mientras que los animales envejecieron.[2]
Representación de un lambeosaurio.En conformación, Lambeosaurus es similar a cualquier otro dinosaurio, pudiéndose mover en dos o en cuatro patas, como lo muestran pisadas de animales relacionados. La cola estaba sostenida por tendones osificados que la mantenían rígida y prevenían que tocara el suelo. Las manos tenían cuatro dedos, faltándole el pulgar de la mano de básica de un tetrápodo, con el segundo tercer y cuarto dedo formando un casco, lo que sugiere que soportaba el peso del cuerpo cuando marchaba. El quinto dedo estaba libre para poder manipular objetos. Cada pie tenia solamente los tres dedos centrales.[3]
La característica más distintiva, la cresta, era diferente en las dos especies mejor conocidas. En L. lambei, tenia forma de hacha cuando el animal estaba completamente crecido, siendo más corta y redondeada en ejemplares considerados hembras.[1] La "hoja del hacha" se proyectaba hacia el frente entre los ojos, y el "mango" era un hueso sólido que salía hacia atrás en la nuca. La " hoja del hacha " tenia dos secciones, la porción superior tenia un hueso delgado, posiblemente sostén de una cresta carnosa que crecería en al llegar a su edad adulta y la porción inferior de la cresta estaba surcada por canales para el aire, continuación de los conductos nasales.[1] En L. magnicristatus, el "mango" estaba muy reducido, y la "hoja" expandida,[4] formando una pomponsa cresta tipo Pompadour. Esta cresta esta dañada en los mejores especímenes, y solamente se conoce la mitad delantera.[5] En la especies menos conocidas L. laticaudus y L. paucidens actualmente la cresta no se conoce. Sin embargo, L. laticaudus puede ser reconocido por su gran tamaño y el perfil alto de su cola, con alargados cheurones y procesos espinosos como los de Hypacrosaurus.[6]
La especie canadiense de Lambeosaurus parece tener un tamaño similar al de Corythosaurus, alrededor de 9,5 metros de largo,[7] pero L. laticaudus era más grande, aproximadamente entre 15 y 16,5 metros de largo y más de 23 toneladas.[6] Impresiones de escamas son conocidas en varios especimenes, estos especimenes son ahora asignados a L. lambei tenía una piel fina y uniforme, con escamas poligonales con una distribución en ninguna sin un orden particular en el cuello, torso, y cola.[8] Un intercalado similar se ve en impresiones asignadas al cuello, brazo y pie L. magnicristatus,[5] considerando que la cola de L. laticaudus con algunos pequeños pedazos óseos encajados entre las escamas hexagonales grandes y redondeadas más pequeñas grandes.[6]
Por medio de sus huellas se ha descubierto que presentaban un comportamiento gregario. Se trataba de animal rápidos, lo que era probablemente su única defensa contra carnívoros, junto con su gran tamaño.
[editar] Historia
Vista de frente de un esqueleto montado de Lambeosaurus lambei en el Museo Tyrrell.Lambeosaurus posee una complicada historia taxonómica, comenzando en 1902 cuando Lawrence Lambe nombro una nueva especie de hadrosáurido (originalmente GSC 419) de Alberta como Trachodon marginatus.[9] Los paleontólogos comenzaron a encontrar mejores restos de hadrosáuridos de las mismas rocas en 1910, en lo que hoy se conoce como la Formación Dinosaur Park del Campaniano Tardío en el Cretácico Superior. Lambe asigno dos nuevos cráneos T. marginatus, y basado en nueva información, acuño el género Stephanosaurus para la especie en 1914.[10] Desafortunadamente, había muy poco para asociar los cráneos al material anterior escaso de marginatus, por eso en 1923 William Parks propone un nuevo género y especie para los cráneos, ambos, el nombre genérico y especific rindiendoloe honores a Lambe: Lambeosaurus lambei (espécimen tipo NMC 2869, originalmente GSC 2869).[11]
[editar] Nuevas especies y procheneosaurios
Aunque los primeros científicos en trabajar en Alberta no lo reconocieran en ese entonces, encontraron los restos de juveniles Lambeosaurus también. Estos fósiles de dinosaurios de pico de pato con cresta de cuerpo pequeño fueron interpretados como adultos de un linaje distinto de hadrosáuridos, la subfamilia Cheneosaurinae.[12] El primero en ser nombrado fue Didanodon altidens, uma mandíbula izquierda (GSC 1092) de la Formación Dinosaur Park descrito por Henry Fairfield Osborn en 1902.[13] Siendo referido a Trachodon como T. altidens por Lambe,[9] y el nombre Didanodon termino rapidamente en la oscuridad no siendo mencionado en el influyente trabajo de Richard Swann Lull y Nelda Wright's d 1942 sobre los hadrosáuridos norteaméricanos.
Procheneosaurus en el Museo Estadounidense de Historia Natural, un posible Lambeosaurus o CorythosaurusEn 1920, William Diller Matthew uso el nombre Procheneosaurus (sin nombre de especie) en una captura pa la fotografia de un esqueleto del for a photograph of a skeleton at the Museo Estadounidense de Historia Natural, de la Formación Dinosaur Park (AMNH 5340).[14] Parks creyó que el procedimiento y la descripción eran inadecuados para que el nombre sea considerado válido, y al tratar la situación, acuñó el género Tetragonosaurus. Dentro de este género el coloco la especie tipo T. praeceps (basandoce en el especímen ROM 3577) y una segunda especie T. erectofrons (ROM 3578) para los pequeños cráneos encontrados en la Formación Dinosaur Park, y asigno al esqueleto de Procheneosaurus de Matthew a T. praeceps.[15] Charles M. Sternberg prosiguio en 1935 adicionando el ligeramente más largo T. cranibrevis, basado en GSC (hoy NMC) 8633.[16] El uso de Tetragonosaurus fue rechazado por Lull en favor de Procheneosaurus. En 1942 junto a Wright transfirieron las especies de Tetragonosaurus y tentativamente a Trachodon altidens, a Procheneosaurus, con P. praeceps sirviendo como especie tipo.[17] Esto se siguió usando hasta 1975, cuando Peter Dodson mostrara que los "cheneosaurios" era realmente individuos juveniles de otros dinosaurios.[1] Procheneosaurus praeceps y altidens son consideardos sinónimos menores de Lambeosaurus lambei,[3] aunque ambos precedan ese nombre.
Los "procheneosaurios" no son los únicos dinosaurios picos de patos crestados que fueron estudiados y nombrados a comienzo del siglo XX. Entonces era la práctica aceptada nombrar géneros y especies para lo qué ahora se ve como más probablemente variación individual, variación debida a la edad o sexo, o distorsión de la fosilización. Tres especies más fueron nombradas durante este período que se relacionan con Lambeosaurus , todos en 1935. Sternberg, en el mismo trabajo del T. cranibrevis , nombro un cráneo y un esqueleto parcial (GSC hoy NMC-8705) L. magnicristatum (corregido a magnicristatus), y un cráneo más pequeño (GSC hoy NMC-8703) L. clavinitialis , con una cresta menos prominente y una espina dorsal reducida en la espalda. PArks contribuyo con Corythosaurus frontalis, basado en el cráneo GSC 5853 (hoy ROM 869), que se diferenció de la bien conocida alta, recta, cresta redondeada de otros especímenes de Corythosaurus teniendo una cresta baja proyectada hacia adelante.[18]
[editar] Consolidación del Género
No hubo descripción de nuevos especimenes desde comienzo de 1900. En 1964 John Ostrom noto que la antigua especie nombrada por Othniel Charles Marsh, Hadrosaurus paucidens, basado en USNM 5457, un maxilar parcial y un escamoso de la Formación Río Judith del Condado Fergus, Montana, sea un espécimen de Lambeosaurus.[19]
En 1975, Peter Dodson, examinando de porqué debe había muchas especies y géneros de dinosaurios de pico de pato lambeosaurinos dentro de un marco de tiempo geológico tan corto y en una pequeña área, publicando los resultados de un estudio morfométrico en el cual midió docenas de cráneos. Encontró que muchas de la especie habían sido basadas en los restos que eran mejor intrepetados como jóvenes o de distintos sexos.Para Lambeosaurus, encontró que L. clavinitialis era probablente un individuo femenino de L. lambei y Corythosaurus frontalis y Procheneosaurus praeceps probablente fueron juveiles de este. L. magnicristatus era bastante diferente autorizar su propia especie. Interpreto a Procheneosaurus cranibrevis y P. erectofrons como coritosaurios jóvenes.[1] Sin embargo, un reestudio de los restos de Procheneosaurus/Tetragonosaurus indican que dentro de la especie, las asignaciones habían llegado a ser confusas, y el tipo espécimen de P. cranibrevis era un Lambeosaurus juvenil, y otros de Corythosaurus, basados en los distintos contactos entre los huesos nasal con el premaxilar.[2]
Restauración en vida de un Lambeosaurus laticaudusPor eso durante los años de 1970, Bill Morris estudio los restos del gigante lambeosaurine de Baja California. Lo llamo L. laticaudus? en 1981 (espécimen tipo LACM 17715). Morris utilizó un signo de interrogación en su trabajo porque no se había encontrado ninguna cresta completa para su especie, y sin ella una asignación definitiva no podría ser hecha. Aunque no se conociera un cráneo, lo consideraba como Lambeosaurus. Interpretó esta especie como acuatica, debido a las características como su tamaño, su cola alta y estrecha (interpretados como una adaptación para nadar), y débil articulaciones de la cadera, así como un hueso del muslo roto y curado, que pensó habría sido demasiada desventaja para que un animal terrestre sobreviva bastante tiempo para curarse.[6]
[editar] Clasificación
Lambeosaurus es el especímen tipo de Lambeosaurinae, una subfamilia de hadrosáuridos que poseían cresta hueca. Como otros lambeosaurinos, esta relacionado cercanamente con Corythosaurus y Hypacrosaurus, diferenciándose por la forma de la cresta.[3] Las relaciones entre estos géneros del dinosaurio son difíciles de realizar. Clasificaciones de Internet suelen incluirlo en la tribu Lambeosaurini, pero esta tribu no esta formalmente definida. Por otro lado, el clado equivalente Corythosaurini ha sido adoptado en publicaciones, incluyéndolo en la ultima filogenia, presentada por David Evans y Robert en su redescripción de L. magnicristatus. Esta revisión encuentra a Lambeosaurus como un taxón hermano a un clado compuesto de Corythosaurus, Hypacrosaurus , y el género ruso Olorotitan. Estos lambeosaurines, con Nipponosaurus , componen a Corythosaurini.[5]
[editar] Especies
Perfiles de varios especimenes de Lambeosaurus, alguna vez asignados a su propia especie, hoy interpretados como individuos de diferentes edades y sexos de L. lambei.Dos especies de Lambeosaurus son actualmente confirmadas como validadas, con una tercera aceptada extensamente y un cuarta aceptada a veces. L. lambei (Parks, 1923) es conocido al menos de 17 individuos, con siete cráneos con material postcraneal y 10 cráneos aislados. L. clavinitialis (C.M. Sternberg, 1935), Corythosaurus frontalis (Parks, 1935) y Procheneosaurus praeceps (Parks, 1931) han sido degradadas a sinónimos de L. lambei en la mayora de las recientes reviciones.[3] Es posibles que los cráneos de L. clavinitialis sin la espina en la nuca podrian representar individuos de L. magnicristatus en lugar de otra,[20] aunque esto fuera rechazada en la redescripción de 2007 de L. magnicristatus.[5] L. magnicristatus (C.M. Sternberg, 1935) se conoce solamente a partir de dos especímenes, ambos con los cráneos. Desafortunadamente, se ha perdido la mayoría del esqueleto articulado del espécimen tipo. Muchos de los huesos estaban dañados extensivamente por el agua mientras durante el almacenaje y desechados antes de la descripción, otras porciones de este esqueleto también se han perdido. Sus restos vienen de rocas levemente más jovenes que L. lambei.[5] El nombre especifico deriva del latín magnus "largo" y cristatus "crestado", refiriéndose a la cresta.[21] Adicionalmente, Jack Horner ha identificado una mandíbula de un lambeosaurine jaws en la Formación Bearpaw de Montana posiblemente perteneciente a L. magnicristatus, este representa al primer lambeosaurino de rocas marinas.[22] El gran L. laticaudus (Morris, 1981), conocido por muchos elementos disociados, de varios individuos, es acxeptado como una potencial especie[3] aunque podria pertenecer también a Hypacrosaurus.[5] [23] Su validez fue discutida por los que describieron Velafrons, quienes encontraron que los restos no son diagnostios pero distintos a los de este género.[23] El nombre proviene del latín latus "amplio" y cauda "cola".[21]
L. paucidens (Marsh, 1889) es degradado a dudoso y enlistado como Hadrosaurus paucidens en los ultimas revisiones,[3] aunque pore lo menos un autor, Donald F. Glut, lo acepta como una especie valida de Lambeosaurus.[24] En este caso, el epíteto específico se deriva del latín pauci- "algunos" y dens "dientes".[21] Las irregularidades de Procheneosaurus cranibrevis y la identidad del tipo como un lambeosaurio juvenil, fue reconocida en 2005,[2] y todavía no se ha incorporado a una circulación amplia. Finalmente, Didanodon y Trachodon altidens han recibido el tratamiento confuso en la revisiones más recientes. Didanodon fue asignado sin comentarios a Lambeosaurus , con todo la especie Trachodon altidens fue enumerado por separado en una tabla de nombres dudosos.[3]
[editar] Paleoecología
Rocas de la Fortmación Dinosaur Park expuesta a lo largo del Río Red Deer, del Parque Provincial del Dinosaurio, en el sur de Alberta.Lambeosaurus lambei yL. magnicristatus, de la Formación Dinosaur Park, fueron miembros de la diversa y bien documentada fauna de animales prehistoricos que incluyen a los ceratopsianos Centrosaurus, Styracosaurus y Chasmosaurus, algunos pico de pato Prosaurolophus, Gryposaurus, Corythosaurus y Parasaurolophus, el tiranosáurido Gorgosaurus, y los anquilosaurianos Edmontonia y Euoplocephalus.[25] La Formación Dinosaur Park se interpreta como una llanura con ríos y terreno aluvional que se convirtia en un pantano y recibía la influencia del cercano Mar Niobraran que trasgredía la porción occidental.[26] El clima era más cálido que el actual de Alberta, sin heladas, pero con temporadas secas y húmedas. Las coníferas formaban parte de la cubierta vegetal dominante, con un sotobosque de helechos, cycas y angiospermas.[27] La similitudes anatómicas de L. lambei, L. magnicristatus y Corythosaurus estaban separados por el tiempo en la misma formación, basado en la estratigrafía. Los fósiles deCorythosaurus en los dos tercios más bajos de la formación, L. lambei están presentes en el tercio superior, y L. magnicristatus solo en la parte más alta, donde la influencia marina era mayor.[28]
[editar] Paleobiología
[editar] Alimentación
Como todo hadrosáurido, los lambeosaurios fueron un herbívoros que alternaban entre un andar cuadrúpedo y bípedo, comiendo plantas con una sofisticado cráneo que le permitía un movimiento análogo a la masticación de los mamíferos. Sus dientes, que se remplazaban continuamente, estaban empaquetados en baterías dentales, que llegaban a contener más de cien dientes, aunque solo un puñado de estos era funcional al mismo tiempo. Usaba un pico para recoger el material vegetal , que era mantenido en la boca por un órgano similar a las mejillas. Se alimentaba de plantas de hasta 4 metros de altura.[3] Como noto Robert Bakker, los lambeosaurinos poseian picos más estrechos que los hadrosaurinos, lo que lleva a pensar que lambeosaurio y sus parientes pudieron ser más selectivos a la hora de comer que estos con sus picos anchos.[29]
[editar] Cresta
Reconstrucción de la alta cresta de L. magnicristatusComo otros lambeosaurinos como parasaurolofo y coritosaurio, un lambeosaurio poseía una distintiva cresta en el techo de la cabeza. La cavidad nasal se continuaba por dentro de la cresta lo que la hacia casi completamente hueca. Mucho se ha sugerido sobre la función o las funciones de la cresta, incluyendo la cubierta de una glándula de sal , una mejora del sentido del olfato, que lo utilizaba como tubo respirador o de aire, actuando como compartimiento de resonancia para hacer sonidos, o como un método para las diversa especies o diversos sexos de la misma especie para reconocerse.[20] [30] Las funciones sociales tales como para la vocalización y reconocimiento se han convertido en lo más extensamente aceptado de las varias hipótesis.[3] El gran tamaño en los hadrosáuridos de la órbita ocular y la presencia de un anillo esclerótico en los ojos implican una visión aguda y de hábitos diurnos, evidencia que la vista era importante para estos animales. El sentido del oído de los hadrosáuridos también parece ser fuerte. Hay por lo menos un ejemplo, en el ' relacionado coritosaurio, de un delgado estribo en el lugar, que combinado con un espacio grande para el tímpano implica un sensible oído medio, y en los hadrosáuridos la lagena es alargada como un cocodriliano. Esto indica que la porción auditiva del oido interno estaba bien desarrollada. [20] Si estuvo utilizada como amplificador de vo, la cresta habría podido también proporcionar las diferencias reconocibles para las diversos especies o sexos, porque las disposiciones de diferenciación de los pasos nasales que corresponden a las diversas formas de la cresta habrían producido sonidos intrínseco diversos. [31]
Como otros paquicefalosáuridos, Pachycephalosaurus era un bípedo posiblemente omnívoro con un techo del cráneo extremadamente grueso. Poseía largas patas y cortos brazos. Pachycephalosaurus es el paquicefalosáurido más grande conocido, midiendo entre 4,5 a 5 metros de longitud y hasta 2000 kilogramos de peso. La bóveda gruesa del cráneo de Pachycephalosaurus y de géneros relacionados dieron lugar a la teoría que los paquicefalosáuridos utilizaron sus cráneos en combate intraespecífico. Esta teoría se ha desacreditado en los últimos años.
La anatomía de Pachycephalosaurus es poco conocida, estando solo su cráneo descrito.[1] Pachycephalosaurus es famoso por tener un gran y huesudo domo en la parte superior del cráneo, de más de 25cm de grosor, lo cuál amortiguó con seguridad su cerebro minúsculo. El aspecto posterior del domo era agudo con nodulos huesudos y los púas huesudas proyectados hacia arriba del hocico. Las puntas eran probablemente redondeados, no agudos.[2]
Comparación de Tamaño entre Pachycephalosaurus wyomingensis y Homo sapiens.El cráneo era corto con las orbitas de los ojo grandes, redondeadas y colocados hacia el frente, sugiriendo que el animal tenía buena visión y era capaz de visión binocular. Pachycephalosaurus tenía un pequeño hocico que terminaba en un pico acentuado. Los dientes eran minúsculos, con las coronas foliformes. La cabeza se apoyaba por un cuello con forma de "S" o "U" -.[2]
Pachycephalosaurus fue probablemente bípedo y uno de los más grandes paquicefalosáuridos conocidos. Comparandolo con otros dinosaurios de cabeza con cúpula mejor conocidos, se puede estimar que Pachycephalosaurus media aproximadamente 4.6 metros de largo,[3] teniendo un cuello bastante corto y grueso, miembros delanteros cortos, un cuerpo robusto, piernas traseras largas y una cola gruesa, que probablemente fue sostenida rígida por tendones osificados.[4]
[editar] Historia
Esqueleto de Pachycephalosaurus exhibido en el Museo Real de Ontario, Toronto.Restos atribuibles a Pachycephalosaurus han sido encontrados desde principios de la década de 1850. Según lo determinado por Donald Baird, en 1859 o 1860, Ferdinand Vandeveer Hayden, un temprano recolector fósiles en el oeste norteamericano, recogió un fragmento del hueso en la vecindad del nacimiento del Río Missouri, qué ahora se reconoce como parte de la Formación Lance en el de sudeste Montana.[5] Este especimén, hoy ANSP 8568, fue descrito por Joseph Leidy en 1872 creyendo que pertenecía a una placa dermal de un reptil o una criatura similar al armadillo.[6] Su naturaleza real no fue revelada hasta que Baird lo volviera a estudiar un siglo más tarde y la identificara como un hueso escamoso (hueso de la parte posterior del cráneo) de Pachycephalosaurus, incluyendo un sistema de nódulos huesudos que correspondían a las encontradas en otros especímenes de Pachycephalosaurus.[5] Debido a que nombre Tylosteus precede a Pachycephalosaurus, de acuerdo con las reglas de ICZN este debería ser el usado. Baird peticiono con éxito que Pachycephalosaurus sea usado en vez del viejo Tylosteus debido a que este último no había sido usado durante los últimos 50 años, basándose en lo no diagnostico del material, y a la pobre información geográfica y estratigráfica.[7] Esto no es el final de la historia, Robert Sullivan sugirió en 2006 que ANSP 8568 es más similar al hueso de un Dracorex que el de Pachycephalosaurus.[1] Este hecho podría revestir actualmente poca importancia, ya que Dracorex es por muchos investigadores considerado un ejemplar juvenil de Pachycephalosaurus, como recientemente ha sido propuesto.[8]
P. wyomingensis, es la especie tipo y única especie valida hasta el momento. Pachycephalosaurus, fue nombrado por Charles W. Gilmore en 1931. El lo acuño para un cráneo USNM 12031, proveniente de la Formación Lance en el Condado de Niobrara, Wyoming. Gilmore lo asigno como una nueva especie de Troodon como T. wyomingensis.[9] En ese tiempo, los paleontólogos solo conocián a Troodon por sus dientes, que eran similares a los de Stegoceras. Por consiguiente, los qué ahora se conocen como paquicefalosáuridos fueron asignados a la familia Troodontidae, una idea incorrecta no corregida hasta 1945, por Charles M. Sternberg.[10]
En 1943, Barnum Brown y Erich Maren Schlaikjer, con nuevo y más completo material establecieron el género Pachycephalosaurus e hicieron a "T. wyomingensis" su especie tipo. También nombraron dos especies mas Pachycephalosaurus grangeri y Pachycephalosaurus reinheimeri. P. grangeri se basa en AMNH 1696, un casi completo cráneo proveniente de la Formación Hell Creek de Ekalaka, Condado Carter, Montana. P. reinheimeri se basaba en lo que es hoy DMNH 469, un domo con algunos restos asociados proveniente de la Formación Lance en el Condado de Corson, Dakota de Sur.[11] Estas otras dos especies fueron posteriormente consideradas Sinónimos de P. wyomingensis por 1983.[12]
En el encuentro anual de la Sociedad de Paleontología de Vertebrados de 2007, Jack Horner de la Universidad Estatal de Montana - Bozeman presento evidencia de que Dracorex y Stygimoloch pueden ser formas juveniles de Pachycephalosaurus.[8]
[editar] Clasificación
Pachycephalosaurus le da el nombre a Pachycephalosauria, un clado de dinosaurios herbívoros ornitisquios que vivieron a finales del período geológico Cretácico, a finales del reinado de los dinosaurios en América del Norte y Asia. A pesar de su andar bípedo, estaban estrechamente más vinculados con los ceratopsianos que con los ornotópodos.[13]
Pachycephalosaurus es el más famos de los miembros dee Pachycephalosauria, a pesar de de no ser el mejor conservado. El clado incluye a Stenopelix, Wannanosaurus, Goyocephale, Stegoceras, Homalocephale, Tylocephale, Sphaerotholus y Prenocephale. Dentro de la tribu Pachycephalosaurini, Pachycephalosaurus esta más estrechamente emparentado con Dracorex y Stygimoloch, aunque estos podrían ser formas inmaduras de Pachycephalosaurus.[8]
[editar] Sistematica
Pachycephalosauria
Stenopelix
Wannanosaurus
unnamed
Goyocephale
unnamed
Homalocephale
Pachycephalosauridae
Stegoceras
Pachycephalosaurinae unnamed
Tylocephale
unnamed
Sphaerotholus
unnamed
Stygimoloch
Pachycephalosaurus
El cladograma presentado sigue el analísi presentado por Williamson & Carr, 2002.[14] Uno de los géneros anteriormente mencionados en el trabajo, Yaverlandia, no es más considerado.[15]
[editar] Paleobiología
Casi todos los fósiles de Pachycephalosaurus de las formaciones Lance y Hell Creek en el oeste de los Estados Unidos.[1] Pachycephalosaurus compartiron territorio con otros parientes cercanos como Dracorex y Stygimoloch.[16] Además había otros dinosaurios en el mismo lugar y tiempo como los ornitópodos Bugenasaura, los hadrosáuridos Edmontosaurus y Anatotitan, ceratópsidos como Triceratops y Torosaurus, el anquilosáurido Ankylosaurus, Y los terópodos Ornithomimus, Dromaeosaurus, y Tyrannosaurus.[17]
Los investigadores alguna vez consideraron a los Pachycephalosaurus y sus parientes eran bípedos similares al muflón o el buey almizclero, en donde los machos compiten entre sí dándose cabezazos. También se creía que su cabeza, cuello, y cuerpo estaban colocados horizontalmente y rectos, para transmitir la tensión durante el choque. Sin embargo, ahora se cree que los paquicefalosaurianos no habrían utilizado sus bóvedas de esta manera.
Cráneo de Pachycephalosaurus.Primera, El techo del cráneo no habría podido soportar adecuadamente el impacto asociado al choque. También, no hay evidencia de las cicatrices o de otro daño en cráneos fosilizado de Pachycephalosaurus.[18] Además, las vértebras dorsales cervicales anteriores muestran que el cuello se llevaba en una posición curvada como una "S" o " U", no con una orientación recta, forma impropia para realizar choque de cabezas directos directo. Por último, la forma redondeada del cráneo disminuiría el área superficial de contacto durante el cabezazo, dando por resultado golpes fallidos.[2]
Lo más probable es que Pachycephalosaurus y los otros paquicefalosáuridos se golpearan mutuamente en los flancos. En este escenario, un individuo pudo haberse colocado paralelo a otro o haberle hecho frente a un rival directamente, usando exhibiciones para la intimidación de su rival. Si la intimidación falla, el Pachycephalosaurus doblaría su cabeza hacia abajo de lado, pegando a su rival en el flanco. Esta hipótesis es apoyada por lo robusto de la mayoría de los paquicefalosáuridos, un rasgo que habría protegido órganos vitales contra el daño. La teoría de los golpes en el flanco fue propuesta primero por Sue en 1978, y ampliado por Ken Carpenter en 1997.[2]
Los paleontólogos no se ponen de acuerdo sobre lo que comía este dinosaurio. Teniendo dientes muy pequeños y surcados, no habrían podido masticar las plantas resistentes y fibrosas con tanta eficacia como otros dinosaurios del mismo período. Se asume que los paquicefalosáuridos vivieron en una dieta mixta de hojas, semillas, frutas e insectos. Sosteniendo esto, los dientes serrados habría sido muy eficaz para destrozar las plantas.[19]
Como es el caso de la mayoría de los dinosaurios, el esqueleto de Parasaurolophus no es completamente conocido. El largo del cuerpo del espécimen tipo de P. walkeri se estima en 10 metros. El cráneo mide 1,6 metros de largo, incluyendo la cresta, sin embargo el cráneo del P. tubicen supera de largo, lo que indica un animal aun más grande.[7] El peso estimado es de 2,5 toneladas.[8] El único miembro delantero conocido es relativamente corto para un hadrosáurido, con una corta pero ancha escápula. El fémur mide 103 centímetros en P. walkeri y es robusto para su longitud cuando es comparado al de los otros hadrosáuridos.[7] El humero y los huesos pelvicos estan fuertemente construidos.[9]
Como otros hadrosáuridos, es posible que caminara tanto en dos como en cuatro patas. Es probable que se parara en cuatro patas para forrajear, pero se desplazaría en dos.[10] El proceso espinoso de las vértebras era alto, algo común en los lambeosaurinos;[7] más alta sobre las caderas, aumentaron la altura de la parte posterior. Las impresiones de la piel que son conocidas en P. walkeri, muestran una estructura uniforme de escamas similares a tubérculos, pero sin ninguna particularidad en especial.[6]
Parasaurolophus walkeri con detalles de la piel.Estos animales tenían la cola larga y aplastada a los lados lo que lleva a pensar que Parasaurolophus podía nadar. Al final de las extremidades es un misterio, algunos paleontólogos dicen que son pezuñas y otros dicen que son garras desgastadas por el tiempo. Se han encontrado impresiones de piel de Parasaurolophus, así como varios esqueletos completos por lo que se tiene una muy buena idea de su aspecto.
[editar] La cresta
Lo más remarcable de este dinosaurio es la cresta, la cuál se despega de la parte posterior de la cabeza y estaba compuesta del hueso premaxilar y el nasal. El especímen tipo de P. walkeri tiene una muesca en las espinas dorsales de las vértebras de cerca donde la cresta golpearía la espalda, pero esto puede ser a patología peculiar a este individuo.[7] William Parks, quien nombrara el género, hipotizo que un ligamento se encontraba desde la cresta a la muesca para ayudar a soportar la cabeza.[6] Aunque esta idea parezca inverosímil,[8] Parasaurolophus es a veces restaurado con una vela de piel desde la cresta al cuello. Esta enorme cresta tubular con cuatro secciones huecas dos que aputaban hacia arriba y dos hacia abajo de largo que se conectaba con la nariz, la cual se creía que podía servir como tubo para respirar debajo del agua, suposición que fue desmentida por estudios posteriores, los cuales establecieron la ausencia de algún tipo de orificio en la parte superior. Se han propuesto muchas hipótesis para explicar la función de la cresta, aunque la mayoría de ellas han sido desacreditadas.[11] [8] Se han dado varias hipotesis acerca de su cresta como respirar debajo del agua teoría que se creyó durante mucho tiempo hasta que fue desmentida. Se cree que era para la atracción y distincion entre machos y hembras, para comunicarse en caso de algun peligro u orden y para termorregulación.[12] La más sobresaliente es la de la comunicación. Esta se realizó observando la estructura interna del tubo, para concluir que pudo funcionar como un resonador natural, y haber tenido una función sonora.[13]
A raíz de modelos hechos de la cresta, se ha sugerido que se empleaba para para emitir sonidos (parecidos a los de un trombón) con la finalidad de comunicarse con dinosaurios de su misma especie. Una función que también se cree posible es la atracción de machos a hembras utilizando la cresta, esto se cree debido a que los machos poseén una cresta más larga que las de las hembras.
La cresta era hueca, con tubos separados que van de cada ventana de la nariz al extremo de la cresta para dar vuelta en la dirección contraria y dirigirse atrás y abajo de la cresta y el cráneo. Los tubos eran más simples en P. walkeri, y más complejos en P. tubicen, donde estaban ocultos y otros tubos cruzados y separados.[14] Como P. walkeri y P. tubicen tenía crestas largas con solamente una curvatura leve, P. cyrtocristatus tenia una cresta corta con un perfil más circular.[15]
[editar] Historia
El especímen tipo de Parasaurolophus walkeri en el Museo Real de Ontario.Fue descrito por primera vez en 1922 por William Parks, se basa ROM 768, un cráneo y esqueleto parcial al que le faltan la mayor parte de la cola, las patas traceras distal de la articulación fémoro-tibio-rotuliana, que fue encontrado por una expedición de la Universidad de Toronto en 1920 cerca de Sand Creek e las riveras del Río Red Deer en Alberta, Canadá.[6] Estas rocas hoy se las data en el Campaniano Cretácico Superior]] siendo parte de la Formación Dinosaur Park.William Parks llamo al especímen P. walkeri en honor a Sir Byron Edmund Walker, Presidente de la junta de administración del Museo Real de Ontario.[6] Los restos de Parasaurolophus son raros en Alberta,[16] con solo otro cráneo de un individuo juvenil, posiblemente de esta misma formación o de la Formación Oldman,[17] y tres posibles especímenes sin cráneos de la Formación Dinosaur Park.[18] En ciertas listas de faunas, se hace mención de material de P. walkeri en la Formación Hell Creek de Montana, una unidad rocosa datada en finales de Maastrichtiano.[19] [20] Esta situación no fue tomada en cuenta por Sullivan y Williamson en su revisión del género en 1999,[14] y no se ha profundizado sobre el tema en otra parte.
En 1921, Charles H. Sternberg recobro un cráneo parcial (PMU.R1250) de la levemente más joven Formación Kirtland en el Condado de San Juan, Nuevo México. Este espécimen se encuentra en Uppsala, Suecia, donde Carl Wiman lo describio como una segunda especie, P. tubicen, en 1931.[21] El epíteto especifico probiene del latín tǔbǐcěn "trompetista".[22] Un segundo cráneo, casi completo P. tubicen (NMMNH P-25100) que fuera encontrado en Nuevo México en 1995. Usando tomografía computada en este cráneo, Robert Sullivan y Thomas Williamson le dieron a este género una monografía muy completa 1999, cubriendo aspectos de su anatomía y taxonomía, y las funciones de su cresta.[14] Williamson publicó posteriormente una revisión independiente de los restos, discrepando con las conclusiones taxonómicas.[23]
John Ostrom describio otro buen ejemplar (FMNH P27393) proveniente de Nuevo México P. cyrtocristatus ein 1961. Incluye un cráneo parcial con una cresta redondeada y corta, y mucho material postcraneal del esqueleto a excepción de los pies, cuello, y de piezas de la cola.[15] El nombre especifico deriva del latín curtus "corto" y cristatus "cresta".[22] Este especimen fue hallado en la Formación Fruitland o, más probable, la base que cubre la formación Kirtland.[14] El rango de distribución de esta especie se amplio en 1979, cuando David B. Weishampel y James A. Jensen describieran un cráneo parcial con una cresta del mismo tipo en el(BYU 2467) from the Campanian-age Kaiparowits Formation of GarfCondado Garfield, Utah.[5] Además de esta, otros cráneo ha sido encontrado en Utah con la cresta de morfología corta y redondeada de P. cyrtocristatus.[14]
[editar] Especies
La especie tipo P. walkeri, de Alberta, es conocida por un solo ejemplar bien establecido.[10] Difiere de P. tubicen by por tener tubos simples en la cresta,[14] y de P. cyrtocristatus por poseer una cresta más larga y no tan redondeada y un humero más largo que el radio.[15]
P. tubicen, de Nuevo México es conocido de por lo menos los restos de tres individuos.[10] Esta es la especie más grande, con pasajes de aires mas complejos en la cresta que P. walkeri y esta en más grande y menos curvada que P. cyrtocristatus.[14]
P. cyrtocristatus, de Nuevo México y Utah, es conocido por tres posibles especimen. Es la especie más pequeña, con una cresta redondeada y corta.[14] Su tamaño pequeño y la forma de su cresta han llevado a varios científicos a sugerir que representa jóvenes o a hembras de P. tubicen, el cuál vivio en el mismo tiempo y de la misma formación. Segun lo dicho por Thomas Williamson, el materia tipo P. cyrtocristatus iaproximadamente del 72% del tamaño de P. tubicen, cerca del tamaño en el cual sobre otros lambeosaurinos se interpreta para comenzar demostrar el dimorfismo sexual en sus crestas (~70% del tamaño del adulto).[23] Esta posición se ha rechazado en revisiones recientes de lambeosaurinos.[10] [24]
[editar] Galería de cráneos
Molde del cráneo de un Parasaurolophus cyrtocristatus, Museo Field.
Molde del Cráneo de unParasaurolophus walkeri skull cast, Museo de Historia Natural de Londres.
Modelo por ordenador de los pasajes dentro de la cresta del cráneo de P. tubicen
[editar] Clasificación
Como su nombre lo indica, Parasaurolophus fue inicialmente vinculado a Saurolophus debido a las similitudes en la cresta.[6] Sin embargo, rápidamente fue asignado a la subfamilia de hadrosáuridos Lambeosaurinae mientras que Saurolophus es considerado un Hadrosaurinae.[25] Es usualmente interpretado de forma separada a otros lambeosaurinos, distinguiéndose de los hadrosáuridos de cresta en forma de casco como Corythosaurus, Hypacrosaurus y Lambeosaurus.[10] [26] [24] Su pariente más cercano parece ser Charonosaurus, un lambeosaurino con un cráneo simila, pero del que no se conoce la cresta completa, de la región del Río Amur en el noreste de China,[27] y los dos son colocado en la tribu Parasaurolophini.[24] P. cyrtocristatus, con su corta y redondeada cresta, es considerado el más filogeneticamente basal de las tres especies conocidas de Parasaurolophus,[24] o podría ser un juvenil o hembra de la especie P. tubicen.[23]
[editar] Paleoecología
Gorgosaurus atacando a P. cyrtocristatus.En la Formación Dinosaur Park, P. walkeri compartía el territorio con una bien documentada fauna de dinosaurios. Los más conocidos de estos son los herbívoros Centrosaurus, Styracosaurus y Chasmosaurus todos ellos dinosaurios con cuernos, Prosaurolophus, Gryposaurus, Corythosaurus, Lambeosaurus que como el Parasaurolophus eran dinosaurios de pico de pato y los dinosaurios acorazados Edmontonia y Euplocephalus. El principal predador fue el tiranosáurido Gorgosaurus, adémas se encontrava el raptor Dromaeosaurus y varios pequeños predadores más.[20] La Formación Dinosaur Park es interpretada como la sección baja de un [río] en una llanura inundable con la presencia de pantanos en influencia marina debido a la Mar interior occidental que invadia las tierras bajas en periodicas ingresiones marinas.[28] El clima era predominantemente cálido, sin estación fría pero si la presencia de temporadas secas y húmedas. Las coníferas eran las plantas predominates en la región, formando la mayor parte de la dosel vegetal. Debajo de este existía un sotobosque de helechos, helechos arbóreos y plantas con flores.[29]
Las especies, P. cyrtocristatus y P. tubicen de Nuevo México compartía territorio con el gran saurópodo Alamosaurus, el hadrosáurido Kritosaurus, el ceratopsiano Pentaceratops, el anquilosáurido Nodocephalosaurus todos ellos hebívoros. Entre los principales carnívoros se encuentra a Saurornitholestes y un gran tiranosáurido que no ha sido nombrado todavía.[20] La Formación Kirtland es interpretada como una planicie ribereña, aparecida despues de que se retirara el Mar Niobraran. Coníferas eran las plantas dominates, y aparentemente los dinosaurios con cuernos erán más comunes que los de pico de pato.[30]
[editar] Paleobiología
[editar] Alimentación
Como todo hadrosáurido Parasaurolophus era un animal con la capacidad de desplazarse en forma bípeda o cuadúpeda, de alimentación herbívora. Tenía cientos de dientes ordenados en columnas que los iba remplazando a medida que se desgastaban. Este desgaste se debía a tenía un sofisticado aparato masticatorio que le permitía moler el alimento antes de tragarlo, a diferencia de otros herbívoros de su época. El aliemnte lo tomaba con un oirgano similar a un pico y era mantenido dentro de la boca por unas estructuras similares a las mejillas de los mamiferos, que impedía que el forraje se le cayera. Este animal comía hasto los cuatro metros del suelo.[10] Segun lo notado Robert Bakker, los lambeosaurinos tenián picos mas estrechos que sus primos los hadrosaurinos, lo que podria implicar que Parasaurolophus y sus parientes pudieron ser más selectivos a la hora de tomar el alimento.[31]
[editar] Cresta craneal
Muchas hipótesis han sido propuestas para explicar las funciones de la cresta craneal del Parasaurolophus, pero muchas han sido refutadas.[8] [32] Actualmente se considera que lo más probale es que haya tenido varias funciones simultáneamente, como parte de un sistema de individualización visualde la especie y sexo, amplificación del sonido para la comunicación y termorregulación.Lo que no está claro es cual fue el principal motor para la evolución de la cresta y de los canales internos.[13]
[editar] Hipótesis rechazadas sobre la función de la cresta
Muchas de las sugerencias tempranas se centraron en las adaptaciones para una forma de vida acuática, siguiendo la hipótesis que los hadrosáuridos eran anfibios, una línea de pensamiento común hasta los años de 1960. Así, Alfred Sherwood Romer propuso que le servia como a tubo respirador,[33] Martin Wilfarth, que era el soporte para una probóscide móvil usada como tubo para respirar o para tomar el alimento,[34] Charles M. Sternberg, que habría servido como trampa de aire para evitar que el agua llegara a los pulmones,[35] y Ned Colbert que habría sido un reservoriao de aire para prolongar su estadía debajo del agua.[36]
Fósil de Parasaurolophus walkeri.Otras sugerencias eran más físicas en naturaleza. Según lo mencionado anteriormente, W. Parks sugirió que se encontraba unida a las vértebras con ligamentos o músculos, y ayudada con el movimiento y soporte de la cabeza.[6] Othenio Abel propuso que era usada como arma para combates con otros de su especie,[37] y Andrew Milner que era usado para abrirse paso entre el follaje, como lo hace el casuario hoy en día.[11] Por otro lado también se propuso que alojaría órganos especiales. Halszka Osmólska sugiere que alojaba una glandula de sal,[38] y John Ostrom que estaba recubierta de tejido sensitivo olfatorio y mejorado sentido del olfato de los lambeosaurinos, que no tenían ninguna capacidad defensiva obvia.[39] Una hipótesis inusual, fue dada por el creacionista Duane Gish, en la que la cresta alojaría un glandulas capases de arrojar un "fuego químico" a sus enemigos, similar a la de los escarabajos bombarderos de hoy en día.[40]
La mayor parte de estas hipótesis se han desacreditado o rechazado.[8] Por ejemplo, no hay agujero en el extremo de la cresta para una función de esnórquel. No hay cicatrices de inserciones musculares para una probóscide y es dudoso que un animal con un pico necesitara una. Como trampa de aire, no habría impedido que entrara el agua. El propuesto depósito de aire habría sido escaso para un animal el tamaño de Parasaurolophus. Otros hadrosáuridos tenían cabezas grandes sin la necesidad de crestas huecas grandes para servir como puntos inserción para los ligamentos de soporte.[39] Ningunas de las propuestas explican porqué la cresta tiene tal forma, porqué otros lambeosaurinos tienen crestas que parecen muy diferentes pero realizan una función similar o cómo los hadrosáuridos sin crestas o con una sólida consiguieron perdurar sin tales capacidades, o porqué algunos hadrosáuridos tenían crestas sólidas. Estas consideraciones afectan particularmente las hipótesis basadas en el aumento de las capacidades de los sistemas ya presentes en el animal, tal como las hipótesis de la glándula de sal y del olfato,[11] e indican que éstas no eran las funciones primarias de la cresta. Además, el trabajo sobre la cavidad nasal de los lambeosaurinos demuestra que nervio olfativo y el tejido sensorial correspondiente estaban en gran parte fuera de la porción de los pasos nasales en la cresta, así que la extensión de la cresta tenía poco que ver con el sentido del olfato.[41]
[editar] Diferencias entre especies, sexo y edad
Diagrama comparativos de las de las crestas craneales de Parasaurolophus cyrtocristatus (a) y Parasaurolophus walkeri (b).Como en otros lambeosaurinos, se cree que la cresta craneal de Parasaurolophus cambiaba con la edad y en el adulto eran diferentes entre los sexos. James Hopson, uno de los primeros investigadores en describir la crestas de los lambeosaurinos en términos de sus diferencias, sugiere que P. cyrtocristatus, con su pequeña cresta es una hembra de P. tubicen.[42] Thomas Williamson en cambio ha sugerido de que se trata de una forma juvenil de este.[23] Ninguna de estas hipotesis ha sido ampliamente aceptada. Con solo seis buenos cráneos y uno juvenil, material adicional es necesario para clarificar esta situación. Williamson noto que en cualquier caso, los jovenes Parasaurolophus probablemente tuvieran, crestas redondeadas como P. cyrtocristatus, que probablemente crecieran rapidamente cuando el animal se hacercara a la madurez sexual.[23] Recientes estudios de cráneos juveniles asignados previamente a Lambeosaurus y hoy considerados Parasaurolophus, provee evidencia de una pequeña cresta tubular presente en los jovenes. This Este espécimen preserva una pequeña marca ascendente en el medio de cada hueso frontal que es similar pero menos notable que las vistas en los adultos. En estos los frontales forman una platforma que soporta la base de la cresta. Este espécimen también indica que el crecimiento de la cresta en Parasaurolophus y el perfil facial de los individuos juveniles se diferenciaba del modelo Corythosaurus-Hypacrosaurus-Lambeosaurus, en parte debido a la cresta de Parasaurolophus careciendo del hueso delgado en forma de peineta que compone la porción superior de la cresta de los otros tres lambeosaurinos.[17]
[editar] Función social
Tomando en cuenta el punto de vista, social y fisiológico se han propuesto las funciones de la cresta, tienen que ver más con la identificación visual y la comunicación sonora. Como un objeto grande, la cresta tiene un claro valor como señal visual y distingue a este animal de sus contemporáneos. El gran tamaño de la órbitas oculares y la presencia de un anillo esclerótico en los ojos implican una visión aguda y hábitos diurnos, evidencia que la vista era importante para estos animales. Si, como se ilustra comúnmente, una vela de piel se extendía desde la cresta al cuello o a la espalda, el efecto de resaltar sobre los demás habría sido incluso más llamativo.[42] Como se ha sugerido por los cráneos de los lambeosaurinos, la cresta de Parasaurolophus le permitiría identificarse de otros dinosaurios similares como Corythosaurus o Lambeosaurus y reconocer el sexo y la edad de otro de su especie por el tamaño y la forma.[13]
[editar] Función en la emisión de sonido
La apariencia externa de la cresta, bastante simple, sin embargo no corresponde a la compleja anatomía interna de los pasajes nasales, lo que sugiere que estos tendrián alguna otra función.[13] Carl Wiman fue el primero en proponer, ya en 1931, que los pasajes habrían servido para emitir un sonido identificatorio, similar a el producido por un cromorno.[21] Hopson & David B. Weishampel revisarón esat idea entre las decadas de 1970 y 1980.[42] [43] [44] Hopson encontró que hay evidencia anatómica que los hadrosáuridos tenían un oído agudo. Hay por lo menos un ejemplo, en el relacionado Corythosaurus, de un delgado estribo (hueso reptil del oído) en el lugar, que combinado con un espacio grande para el tímpano implica un sensible oído medio. Además, en los hadrosáuridos el extremo del ducto coclear es alargado como en los cocodrilianos, indicando que la porción auditiva dle oído interno estaba bien desarrollada.[42] Weishampel suguiere que P. walkeri es capaz de producir frecuencias de sonido de entre 48 a 240 Hz, y P. cyrtocristatus (interpretada como una cresta de forma juvenil) de entre 75 a 375 Hz. Basados en la similitud de los oídos internos de los hadrosáuridos con los de los cocodrilos, propuso que los adultos eran sensibles a las frecuencias altas, como las que producirían las crías. According to Weishampel, this is consistent with parents and offspring communicating.[43]
P. cyrtocristatus en el Museo Field.Un modelo de ordenador de la cresta bien preservada de P. tubicen, que poseía pasajes de aires más complejos que P. walkeri, llevaron a la reconstrucción de los posibles sonidos que produciría.[45] La trayectoria principal resuena aproximadamente 30 Hz, pero la anatomía complicada de los senos causa picos y los valles en el sonido .[46]
[editar] Función en la termorregulación
El gran areas expuesta y la vascularización de la cresta ha llevado a pensar que también tendría función en la termorregulación.[47] P.E. Wheeler fue el primero en proponer este uso en 1978 como una manera de enfriar el cerebro.[48] Teresa Maryańska y Osmólska propusieron la función termorregulatoria casi al mismo tiempo,[49] y Sullivan & Williamson lo tomaron como objeto de estudio. La disertación de David Evans de 2006 sobre la función de la cresta de los lambeosaurinos, por lo menos como factor inicial para la evolución de la extensión de la cresta.[13]
Iguanodon fue un robusto herbívoro que podía alternar entre la posición bípeda a cuadrúpeda.[1] La especie bien conocida, I. bernissartensis, se ha estimado que pesaba alrededor de 3 toneladas en promedio,[2] y a medir alrededor de 10 metros de largo cuando adulto, con algunos especímenes que posiblemente habrían alcanzado los 13 metros.[3] Otras especies no eran tan grandes, el igualmente robusto; I. dawsoni se estima en 8 metros de largo, y su contemporáneo ligeramente construido, I. fittoni en 6 metros.[4] Este género tenía un cráneo grande, alto pero estrecho, con un desdentado pico cubierto probablemente con queratina, y los dientes como los de una iguana, pero mucho más grande y juntos en paquetes.[1]
Los brazos eran largos, hasta 75% de la longitud de las piernas en I. bernissartensis y robusto,[3] con las manos poco flexibles construidas de modo que los tres dedos centrales pudieran soportar el peso.[1] Los pulgares eran los púas cónicas que se volteaban hacia fuera lejos de los tres dedos principales. En restauraciones tempranas, la púa fue puesta en el la nariz del animal. Fósiles posteriores revelaron la naturaleza verdadera de las púas del pulgar,[5] estando su verdadera función en debate. Habrían podido ser utilizados para la defensa, o para forrajear el alimento. El dedo meñique era alargado y diestro, y habría podido ser utilizado para manipular objetos. Las piernas eran poderosas, pero no construido para correr, y tenía tres dedos en cada pie. La espina dorsal y la cola fueron apoyadas y atiesadas por tendónes osificados, que eran los tendones que se volvieron hueso durante vida, estos huesos como cilindros se omiten generalmente de los montajes y de los dibujos esqueléticos.[1] En general, su aspecto y estructura del cuerpo no difería de los posteriores hadrosáuridos.
Estos ornitópodos, así como podían caminar en cuatro patas, también podían adoptar una postura bípeda ya que la columna vertebral se sostenía horizontalmente acercando los brazos al suelo, de manera que los iguanodontes pudieran lograr tal habilidad. La cola también estaba en posición horizontal alejada del cuerpo y por encima del suelo. En un comienzo, los científicos pensaron que los iguanodontes eran animales torpes, como un rinoceronte, al caminar en cuatro patas. Más tarde, cuando en Bélgica se encontraron ciertos esqueletos, creyeron erróneamente que se desplazaban erguidos con la cola apoyada en el suelo. Sus brazos eran largos y sus dedos se doblaban hacía atrás para sostener el cuerpo al caminar en cuatro patas. El quinto dedo era delgado y flexible.
[editar] Historia
Ilustración de los fósiles de dientes de Iguanodon junto a los de la moderna iguana en el trabajo de Mantell de 1825 sobre Iguanodon.
Restos encontrados en Maidstone en 1840.El descubrimiento de Iguanodon ha sido acompañado de largo del tiempo por una popular leyenda. El primer rastro de un Iguanodon fue un diente fosilizado encontrado por la esposa de Gideon Mantell, Mary Ann Mantell, en los estratos del Bosque Tilgate, Cuckfield, Inglaterra, mientras su marido revisaba un paciente. Sin embargo, no hay evidencia que Mantell llevara a su esposa con mientras veía a sus pacientes. Además, admitió en 1851 que él sí mismo había encontrado los dientes.[6] Ninguno afirma que la otra historia es falsa[7] Sin importar las circunstancias exactas, inpecciono el área en busca de más fósiles, y consultó a los expertos sobre fósiles de su tiempo en cuanto a qué clase de animal pudieron pertenecer los huesos. La mayor parte de los científicos, por ejemplo William Buckland y Georges Cuvier, pensaron que los dientes eran un pez o mamífero. Sin embargo, Samuel Stutchbury, un naturalista del colegio real de cirujanos, reconocido que se asemejaban a los de una iguana, no obstante veinte veces de más grande.[2] Mantell no describio sus descubrimientos hasta 1825, cuando presentara un trabajo en la Royal Society de Londres.[8] [6] En el reconocimiento de la semejanza de los dientes a los de la iguana, Mantell nombró su nuevo género Iguanodon o "dientes de iguana", a partir de iguana y del griego odontos ("diente").[3] Basandose en una escala isometrica, estimo que la criatura habría alcanzado los 12 metros de largo.[8] La idea inicial fue llamarlo Iguanasaurus ("lagarto iguana "), pero su amigo William Daniel Conybeare le sugirió que ese nombre era más aplicable a la misma iguana y que mejor seria llamarlo Iguanoides ("como una iguana ") o Iguanodon.[9] [10] Tuvo el descuido de no asignar nombre a la especie para formar un apropiada nomenclatura binomial, así que uno le fue suministrado en 1829 por Friedrich Holl: I. anglicum , que fue enmendada más adelante a I. anglicus.[11]
Restauración de Mantell de Iguanodon basado en los restos de Maidstone.Años después se encontró un espécimen más completo de Iguanodon en una cantera de Maidstone, el que Mantell adquirió e identificó basado en el diente que tenía. La losa de Maidstone permitió las primeras reconstrucciones esqueléticas y las interpretaciones artísticas de un iguanodonte. El error más famoso fue la adición de un cuerno, también descubierto por la esposa de Mantell, sobre la nariz del dinosaurio.[12] El descubrimiento de muchos mejores especímenes de I. bernissartensis en 1878 reveló que el cuerno era en realidad un pulgar modificado, quizás usado para la defensa, a modo de puñal contra los depredadores de su ambiente. Inclusive, dicho error se encuentra aún presente en las esculturas inauguradas en la exposición del Palacio de Cristal, conservadas actualmente como testimonio histórico. Para conmemorar el descubrimiento incorporo un Iguanodon como tenantes en su escudo de armas en 1949.[13] Este espécimen se ha ligado al nombre I. mantelli , una especie nombrada en 1832 por Christian Erich Hermann von Meyer en lugar de I. anglicus , solamente porque viene realmente de una diferente formación geológica rocosa que el material original I. mantelli / I. anglicus.[10]
El famoso banquete de Benjamin Waterhouse Hawkins dentro del Iguanodon.Al mismo tiempo, la tensión comenzó a crecer entre Mantell y Richard Owen, un científico ambicioso con una financiación mucho mejor y las conexiones de la sociedad en los mundos turbulentos del acto de la reforma de 1832. Owen, una firme creacionista, se opuso a las versiones tempranas de ciencia evolutiva ("transmutacionismo") que entonces estaba siendo discutida y utilizo el termino que el mismo acuñaría como dinosaurios como arma en este conflicto. Con la descripción de Dinosauria redujo proporcionalmente las longitudes de los dinosaurios de los excesivo 61; metros, determinado que no eran simplemente lagartos gigantes, y propuso que eran avanzados como los mamíferos, características dadas a ellos por Dios, según la comprensión de su tiempo, no habrían podido ser " transmutado" de reptiles a las criaturas como mamífero.[14] [15]
Poco antes su muerte en 1852, Mantell argumentó que Iguanodon no era un pesado animal similar a un paquidermo[16] como Owen proponía, pero no fue invitado a participar en la creación de los dinosaurios de The Crystal Palace, que que coloco la imagen de los dinosaurios de Owen en la imaginación popular.[14] Junto a Benjamin Waterhouse Hawkins, creo casi dos docenas de esculturas de tamaño natural de los varios animales prehistóricos construidos enconcreto sobre un marco de acero y ladrillo; dos Iguanodon , uno parado y el otro sobre su vientre, estaban incluidas. Antes que la escultura de pie de Iguanodon fuera terminada, se llevó a cabo un banquete en su interior para veinte comensales.[17] [18] [19]
[editar] La Mina de Bernissart
Fotografía de un esqueleto de Iguanodon de Bernissart siendo montado.El más grande descubrimiento de restos de Iguanodon ocurrió en 1878 en la mina de carbón de Bernissart en Bélgica, a una profundidad de 322 metros.[5] Con el estímulo de Alphonse Briart; el supervisor de minas de Morlanwelz, Louis Dollo, con Louis de Pauw, supervisó la excavación de los esqueletos y los reconstruyó. Se recogieron al menos 38 Iguanodon individuales,[1] la mayoría de ellos adultos.[20] Muchos de ellos colocados en exposición publica en 1882 donde todavía presente están 11 son montajes exhibidos erguidos, y 20 como aproximadamente fueron encontrados.[5] Es la exhibición más impresionante en el Real Instituto Belga de Ciencias Naturales, en Bruselas. Una replica de estos es exhibida en Museo de Hstoria Natural de la Universidad de Oxford y en el Museo Sedgwick de la Ciencias de la Naturaleza en Cambridge. La mayoría de los especímenes están referidos a una nueva especie, I. bernissartensis, un animal más grande y pesado que los restos encontrados en Inglaterra, pero un espécimen fue referido el nebuloso y grácil I. mantelli (hoy Dollodon bampingi). Los esqueletos eran algunos de los primeros esqueletos completos del dinosaurio conocidos. Fueron encontrados con los esqueletos los restos de plantas, peces, y de otros reptiles,[5] incluyendo el cocodriliano Bernissartia.
Iguanodon bernissartensis Dibujos de los fósiles de 1882.La ciencia de la conservación de los restos fósiles estaba en su infancia, y estaban mal equipados para tratar los que se conocía como "la enfermedad de la pirita". La pirita en los huesos cambiaba a sulfato del hierro, dañando los restos haciéndolos agrietarse y desmenuzándolos. Cuando estaban en la mina, los huesos fueron expuestos a la humedad que evitó que sucediera esto, pero cuando estaba fueron expuestos al aire abierto más seco, la conversión química natural comenzó a ocurrir. Al no saber la causa verdadera, y el pensamiento de ella eran una infección real, el personal en el museo en Bruselas frustrada para tratar el problema con una combinación de alcohol, arsénico, y laca. Esta combinación fue pensada para penetrar (alcohol), para matar cualquier agente biológico (arsénico), y para endurecer simultáneamente (laca) los fósiles. Este tratamiento tenía el efecto involuntario del aislamiento en humedad y de prolongar el período de daño. Los tratamientos modernos en lugar de esto implican la supervisión de la humedad del almacenaje del fósil, o, para los especímenes frescos, preparando una capa especial de polietilenglicol que entonces se calienta en una bomba de vacío, así que la humedad se quita inmediatamente y el espacio de poro se infiltra con polietilenglicol para sellar y consolidar el fósil.[5]
Uno de los varios esqueletos mostrados en Bruselas.Los especímenes de Dollo permitieron que se demostrara que los paquidermos prehistóricos de Owen no eran la imagen correcta de Iguanodon. En su lugar modeló los montajes esqueléticos basándose en el emú y el walabí, y puso la púa que había estado en la nariz firmemente en el pulgar.[21] [22] No estaba totalmente correcto, sino que también ya que tenía la desventaja de haber frente al primer dinosaurio completo. Un problema que fue reconocido más adelante era la curva que introdujo en la cola. Este órgano era más o menos recto, como se muestra por los esqueletos que él excavaba, y la presencia de tendones osificados. De hecho, para conseguir la curva en la cola para una más parecida a la del walabí o canguro, la cola habría tenido que estar rota. Con su cola y parte posterior correctas, rectas, el animal habría caminado con su cuerpo horizontal a la tierra, con los brazos para apoyar el cuerpo si fuera necesario.[5] Las excavaciones en la mina pararon en1881, aunque no fuera agotada de fósiles, como reciente las operaciones de la perforación han demostrado.[23] Durante la Primera Guerra Mundial, cuando la ciudad fue ocupada por los alemanes, preparaciones fueron hechas para abrir de nuevo la mina para la paleontología y Otto Jaekel fue enviado desde Berlín para supervisarlas. Cuando los aliados recuperaron Bernissart apenas la primera capa fosilífera estaba a punto de ser destapada. Otras tentativas de abrir de nuevo la mina fueron obstaculizadas por problemas financieros y paradas en conjunto en 1921 en que la mina se inundó.[5]
[editar] Últimos descubrimientos en todo el mundo
La investigación sobre Iguanodon disminuyo durante la primera parte del siglo XX cuando las Guerra Mundiales y la Gran Depresión envolviendo a Europa. Una nueva especie que se convirtió en el tema de mucho estudio y de controversia taxonómica , I. atherfieldensis, así llamada en 1925 por R. W. Hooley, para un espécimen recolectado en Atherfield Point en la Isla de Wight.[24] Sin embargo, el cuál había sido un género europeo ahora era encontrado en todo el mundo, con el material en África (dientes en Túnez,[25] y en otras partes del Desierto del Sahara),[26] Mongolia (I. orientalis),[27] y los Estados Unidos en América del Norte (I. ottingeri de Utah[28] ). Otra especie de norteamérica, de Dakota del Sur, han sido asignadas alguna vez a Iguanodon como I. lakotaensis,[29] siendo reclasificada como Dakotadon.[30]
Iguanodon no fue parte del principio del renacimiento de los dinosaurios que comenzó con la descripción del Deinonychus en 1969, pero no fue olvidado por mucho tiempo. El trabajo sobre mecanismos de alimentación de los ornitópodos de David B. Weishampel proporcionó una mejor comprensión de cómo alimentó,[31] y David B. Norman trabajo con muchos aspectos de Iguanodon, haciéndolo uno de los dinosaurios más conocidos del mundo[32] [5] [33] [1] Ademas de el hallazgo de numerosos esqueletos de Iguanodon, en Nehden, Renania del Norte-Westfalia, Alemania, proveyó la evidencia de la vida en manada en este género ya que los animales en el área del hallazgo parecen que habian estado cerca cuando una inundación repentina los mato. Al menos 15 individuos, a partir del 2 metros de largo, se han encontrado alli, aunque por lo menos algunos de ellos sean iguanodóntidos gráciles y pertenezcan al relacionado Mantellisaurus o Dollodon (descrito como I. atherfieldensis , en por creerse que se trataba de otra especie de Iguanodon).[20] [30]
Material de Iguanodon fue usado en la busqueda de ADN y otras biomoleculas. En el estudio de Graham Embery et al, huesos de Iguanodon fueron prosesados en busquedad deproteinas. En esta investigacióin, restos identificables de tipicas proteinas de hueso, como fosfoproteinas y proteoglicanos, fueron encontrados en una costilla.[34]
[editar] Clasificación
Cladograma simplificado de Iguanodontia, creado después de Norman (2004).[1]Iguanodon le da al nombre al clado sin clasificar Iguanodontia, un grupo muy populoso de ornitopódos con muchas especies conocidas del Jurásico Medio al Cretácico Superior. De este grupo Iguanodon es el mejor conocido, en el cual se incluyen también Dryosaurus, Camptosaurus, Ouranosaurus, y los dinosaurios pico de pato o hadrosáuridos. En fuentes viejas, Iguanodontidae fue mostrada como una familia distinta.[35] [36] Esta fasmilia tradicionalmente ha sido usada como un taxón cajón de sastre, incluyendo ornitopódos que no eran hipsilofodóntidos o hadrosáuridos. En la práctica, animales como Callovosaurus, Camptosaurus, Craspedodon, Kangnasaurus, Mochlodon, Muttaburrasaurus, Ouranosaurus, y Probactrosaurus fueron usualmente asignados a esta familia.[36] Con el advenimiento del análisis cladistico, Iguanodontidae como era interpretado tradicionalmente de demostró que era parafilético, y estos animales se colocan en diversos puntos refiriéndolos a los hadrosáuridos en un cladograma, en vez de en un solo clado distinto.[37] [1] Esencialmente, el concepto moderno de Iguanodontidae incluye actualmente solamente Iguanodon. Los grupos como Iguanodontoidea todavía se utilizan como clados sin clasificar en la literatura científica, aunque muchos iguanodóntidos tradicionales ahora se incluyen en la superfamilia Hadrosauroidea. Iguanodon es ubicado entre Camptosaurus y Ouranosaurus en cladogramas, y se desciende probablemente de a animal similar al camptosaurio. [1] En este punto, Jack Horner sugiere, basado sobre todo en las características del cráneo, que debido a que los hadrosáuridos formaron realmente a dos grupos distantemente relacionados, con Iguanodon en la línea de los de las cabezas chata hadrosaurinos, y Ouranosaurus en la línea de los crestados lambeosaurinos, [38] pero esta propuesta ha sido rechazada.[37] [1]
[editar] Especies
Iguanodon bernissartensis montado correctamente en su postura de cuadrúpedo, Instituto Real de Ciencias naturales de Bélgica Bruselas.Debido a que el iguanodonte es uno de los primeros géneros de dinosaurios en haber recibido un nombre, son numerosas las especies que han sido clasificadas allí. Si bien el iguanodonte nunca ha sido un Taxón cajón de sastre (al estilo de otros géneros de dinosaurios como los Megalosaurus y los Pelorosaurus) su historia ha sido complicada y su taxonomía debe ser revisada constantemente.[39] [40] [41] [30] Los restos de las especies más conocidas han surgido en Bélgica, Inglaterra, España y Francia. Restos de animales similares, posiblemente pertenecientes a este género han sido encontrados también en Túnez y Mongolia, y de especies diferenciadas en Utah, Estados Unidos. Gregory Paul ha recomendado limitar el uso de I. Bernissartensis estrictamente a los descubrimientos de Bernissart, y utilizar I. sp. (que significa especie indeterminada) para los restos de robustos animales tipo iguanodonte en rocas europeas del Barremiano.[30] Al final, lo que inicialmente se pensó que era un dinosaurio típico de las islas Británicas puede en realidad haber escasamente habitado allí.
I. Anglicus fue la especie tipo original, pero el holotipo estaba basado en un solo diente y desde entonces sólo se han encontrado restos parciales de la especie. En marzo de 2000, la Comisión Internacional de Nomenclatura Zoológica cambió la especie tipo a la más conocida I. Bernissartensis. El diente original del iguanodonte es guardado en el Te Papa Tongarewa, el museo nacional de Nueva Zelanda en Wellington, aunque no está exhibida actualmente. EL fósil llegó a Nueva Zelanda luego que el hijo de Gideon Mantell se estableciera allí.[42]
[editar] Especies actualmente validadas
Sólo unas pocas de las especies originalmente asignadas a la familias de los iguanodóntidos son actualmente consideradas válidas, y de ellas, sólo una corresponde al género Iguanodon.[1] [30] I. bernissartensis, es descrito por George Albert Boulenger en 1881, siendo el neotipo para el género. Esta especie es la más conocida para los muchos esqueletos descubiertos en Bernissart, pero también se conoce de los restos a través de Europa. David Norman sugiere que se incluya a la dudosa I. orientalis de Mongolia,[43] pero esto no ha sido seguida por otros investigadores.[30]
Dos especies descritas por Richard Lydekker a finales del siglo XIX son válidas, pero raramente mencionadas. I. Dawsoni, descrita por Lydekker en 1889,[44] es conocida por dos esqueletos parciales encontrados en Sussex del Este, Inglaterra,[1] de mitades del Valanginiano durante el Cretácico inferior.[30] I. Fittoni fue tanbién descrito por Lydekker, en 1888.[45] Como el I. Dawsoni, esta especie también proviene de la Formación Arcilla de Wadhurst[30] de Sussex del Este. Restos encontrados en España podrían también pertenecer al género. Las dos especies se diferencian por en base a características en la pelvis y en las vértebras, en tamaño y en contextura.[4] Por ejemplo, I. Dawsoni era más robusto que I. Fittoni, con grandes vértebras al estilo del Camptosaurus y una columna vertebral corta, mientras que el I. Fittoni es conocido por su columna vertebral larga, estrecha e inclinada.[1] Sin embargo, su pertenencia al género podría ser cuestionada en un futuro próximo.[30]
[editar] Especies reasignadas
Dos especies de iguanodonte propuestas por Richard Owen han sido reasignadas a otros géneros. I. hoggi (también llamada I. boggii o hoggii), bautizada por Owen en base a una mandíbula inferior hayada en la Formación Purbeck del Titoniano-Berriasiano Jurásico superior-Cretácico inferior del Lecho Purbeck en Dorset en 1874, fue reasignada como Camptosaurio por David Norman y Paul Barrett,[40] aunque esta reasignación es actualmente cuestionada.[30] [46] I. major, bautizada a partir de una vértebra encontrada en la Isla de Wight descrita por Owen en 1842 como una especie de Streptospondylus, es todavía un nomen dubium que puede ser un sinónimo de I. anglicus,[1] o una especie independiente.[47]
Adicionalmente, otras catorce especies consideradas originalmente como iguanodontes han sido reclasificadas en otros géneros. La especie Iguanodon albinus (o Albisaurus scutifer), descrita por el paleontólogo checo Antonin Fritsch (Frič en su ortografía original) en 1893, es un reptil dubious nondinosaurian conocido como Albisaurus albinus.[48] I. atherfieldensis, descrita por Reginald Hooley en 1925,[24] era más pequeño y menos robusto que I. Bernissartensis, con una columna vertebral más larga. Fue renombrada como Mantellisaurus atherfieldensis en 2007.[41] I. exogyrarum (también escrita como I. exogirarum o I. Exogirarus) fue descrita por Fritsch en 1878. Es un nomen dubium en base al poco material que existe de ella y fue reasignada por George Olshevsky, como Ponerosteus.[49] I. Valdensis, descrita por Hulke en 1879 a partir de restos de vértebras y de la pelvis, encontrados en la Isla de Wight en una capa del Barremiano,[50] originalmente llamada Vectisaurus, puede ser un espécimen de Mantellisaurus atherfieldensis que no llegó a edad adulta,[51] o de una especie no determinada de Mantellisaurus.[30] I. Gracilis, bautizada por Lydekker en 1888 como especie tipo de Sphenospondylus y asignada como iguanodonte en 1969 por Rodney Steel, podría también pertenecer a Mantellisaurus atherfieldensis.[1]
I. foxii (también escrita I. foxi) fue descrita originalmente por Thomas Henry Huxley en 1869 como especie tipo de Hypsilophodon; Owen (1873 o 1874) la reasignó a los iguanodontes, pero su asignación fue rápidamente contestada.[52] I. hollingtoniensis (también conocida como I. Hollingtonensis), descrita por Lydekker en 1889, ha sido considerada como sinónimo de I. fittoni,[1] . Orto espécimen asignado a I. hollingtonensis por Richard Owen en 1874, con una combinación inusual de mandíbula inferior al estilo hadrosaurid y un antepecho robusto, puede representar un taxón no clasificado.[30] I. prestwichii (también escrito I. Prestwichi), desrito por John Hulke en 1880, fue reasignado como Camptosaurus prestwichii. I. seeleyi (escrito igualmente I. seelyi), descrito por Hulke dos años después del I. prestwichii, has sido considerado sinónimo de I. bernissartensis,[1] aunque todavía esto sigue en disputa.[30] [47] I. suessii, descrito por Emanuel Bunzel en 1871, fue reasignado como Mochlodon suessi.[1]
I. lakotaensis fue descrito por David B. Weishampel y Philip R. Bjork en 1989.[29] Es la única especie de iguanodonte de América del Norte ampliamente aceptada, I. Lakotaensis fue descrito a partir de un cráneo parcial encontrado en la formación Lakota de Dakota del Sur, EE.UU. del período Barremiano durante el Cretácico inferior. Su asignación ha generado controversia. Algunos investigadores sugieren que es más basal que el I. Bernissartensis, y relacionado con el género Theiophytalia,[53] pero David Norman ha sugerido que puede ser un sinónimo de I. bernissartensis.[39] Posteriormente, Gregory S. Paul le asignó su propio género, Dakotadon.[30]
IRSNB 1551, el holotipo de Dollodon bampingi, anteriormente Iguanodon mantelli, mostrado en el Instiuto Real Belga para las Ciencias Naturales en Bruselas.Iguanodon mantelli (también escrito I. manteli o I. mantellii), descrito por Christian Erich Hermann von Meyer en 1832, está realmente basado en el mismo material que I. anglicus.[54] Sin embargo, algunos esqueletos, incluyendo el espécimen Maidstone y uno de loas especímenes de esqueletos Bernissart han sido por años asignados aquí, y su atribución no ha sido completada. El esqueleto grácil Bernissart, por ejemplo, fue reasignado, primero como Mantellisaurus atherfieldensis,[33] y, luego de una revisión más exhaustiva, a su propio gpenero y especie, Dollodon bampingi.[30] I. orientalis, descrito por A. K. Rozhdestvensky en 1952,[27] se basó en muy poco material, pero un cráneo y un hocico arqueado que le estaban asignados fueron reclasificados como pertenecientes a un Altirhinus kurzanovi en 1998.[39] Al mismo tiempo, I. orientalis se consideró como un nomen dubium no distinguible de I. bernissartensis.[43] [39] Harry Seeley describió a I. phillipsi en 1869,[55] pero posteriormente lo reasignó a Priodontognathus.[56]
[editar] Especies dudosas
Cinco especies de iguanodonte son consideradas como nomina dubia o no descritas. I. anglicus, descrita por Friedrich Holl en 1829,[11] es la especie tipo original de Iguanodon, pero, como se mencionó antes, pue remplazada por I. Bernissartensis. En el pasado ha sido escrita como I. angelicus (Lessem y Glut, 1993) y I. anglicum (Holl, 1829 corregido, Bronn, 1850). Es conocida por las dientes encontrados en el bosque de Tilgate de Sussex del Este, Inglaterra del Valanginiano-Barremiano, del Cretácico inferior. I. hillii, bautizada por Edwin Tully Newton en 1892 en base a un diente encontrado en la formación Lower Chalk de Hertfordshire del Cenomaniano, parte del Cretácico superior, es un miembro de los hadrosáuridos.[57] Los restos de un "I. mongolensis" (Whitfield, 1992, nomen nudum de un comentario en la foto de un libro) fueron posteriormente nombrados Altirhinus.[58]
I. ottingeri, descrito por Peter Galton y James A. Jensen en 1979, es un nomen dubium basado en dientes encontrados en la porción infeior de la Formación Montaña Cedar de Utah, probablemente del Aptiano.[28] I. praecursor (también escrito I. Precursor), descrito por E. Sauvage en 1876 a partir de dedos from dientes de una formación en Pas-de-Calais, Francia, es actualmente un saurópodo, que a veces ha sido asignado clasificado como Neosodon,[59] Si bien estos dos vienen de formaciones diferentes.[60]
Finalmente, otros géneros y especies poco conocidos están incluidos entre los Iguanodon sin ser especies separadas, aunque su asignación podría cuestionarse con la la reasignación de I. atherfieldensis. Estos incluyen Heterosaurus neocomiensis (Cornuel, 1850), Hikanodon (Keferstein, 1834), y Therosaurus (Fitzinger, 1840), y las especies "Streptospondylus" recentior (Owen, 1851), "Cetiosaurus" brachyurus,[2] y parte de "C." brevis (Owen, 1842; "C." brevis es una quimera paleontológica).[61] El nomen nudum "Proiguanodon" (van den Broeck, 1900) también está en este grupo,[62] y posiblemente también los poco conocidos "Streptospondylus" grandis (Owen, 1851) y meyeri (Owen, 1854).[10]
[editar] Paleobiología
[editar] Alimentación y dieta
Cranéo de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.Uno de los primeros detalles notados en los iguanodóntes fue que tenia dientes de reptil herbívoro,[8] aunque no haya habido siempre consenso en cómo comió. El mismo Mantell noto que los restos con los que trabajaba eran distintos a lo de cualquier reptil moderno, especialmente en la desdentada mandíbula inferior con una sínfisis de forma de cuchara, que comparo con el perezoso de dos dedos y el extinto perezoso terrestre Mylodon. También sugirió que el iguanodontetenía una lengua prensil que se podría utilizar para recolectar el alimento,[63] como la jirafa. Restos más completos han demostrado que esto era un error; por ejemplo, el hueso hiodeo que soportaba la se construye pesadamente, implicando una lengua muscular, no prensil usada para mover el alimento alrededor en la boca. [32] The giraffe-tongue idea has also been incorrectly attributed to Dollo via a broken lower jaw.[64]
Los dientes de los iguanodontes, como el nombre sugiere, son similares a los de la iguana, pero mayores. A diferencia que los hadrosáuridos, que tenían columnas de reemplazos de dientes, los iguanodontes tenían un solo reemplazo del diente por vez. El maxilar soportada a 29 dientes por lado, con ningunos en los premaxilares, y el dentario 25, los números son diferentesd porque los dientes en la mandíbula inferior son más amplios que ésos en la parte superior.[37] Porque las filas del diente tienen una profunda inserción en la parte oral de mas quijadas, y debido a otros detalles anatómicos, se cree que, como con la mayoría de los otros ornitisquios, los iguanodontes tenía una clase de estructura como la mejilla muscular o no-muscular, para conservar el alimento en la boca.[65] [66]
Una mano de Iguanodon mostrada en Bruselss; con el dedo prensil extendido.El cráneo fue estructurado de una manera tal que cuando cerraba la boca, los huesos que sostienen los dientes en la quijada superior se arquearan hacia fuera. Esto causaría las superficies más bajas de los dientes de la quijada superior frotaran contra la superficie superior de los dientes de la mandíbula inferior, moliendo cualquier cosa entre ellos mientras tanto y proporcionando una acción que es el equivalente al la masticación de los mamíferos.[31] Debido a que los dientes fueron substituidos siempre, el animal habría podido utilizar este mecanismo a través de su vida, y podría comer material vegetal resistente.[67] Adicionalmente, las partes frontales de las mandíbulas del animal de eran desdentadas e inclinadas con nodos huesudos, superiores e inferiores,[1] proporcionando un margen áspero que fue cubierto y alargado probablemente por el material tipo queratina para formar un pico para arrancar con los dientes ramitas y retoños.[5] La recolección del alimento habría sido ayudada por su flexible dedo menor, que se habría podido utilizar para manipular objetos, a diferencia de los otros dedos.[1]
Exactamente qué comian los iguanodontes con sus quijadas bien desarrolladas no se sabe. El tamaño de la especie más grande, por ejemplo I. bernissartensis, pudo haberlo ayudado a alcanzar el follage de árboles a 4 o 5 metros de alto.[3] Una dieta de equisetos, cycas y coníferas fue sugerida por David Norman,[5] aunque los iguanodontídos en general se hayan comido angiosperma en el cretácico el debido al los hábitos deducidos del ramoreo bajo. El crecimiento de las angioespermas, según esto hipótesis, habría sido animado por los iguanodontesque alimenta porque las gimnospermas serían quitadas, permitiendo que más espacio para la mala hierba - como las primeras angiospermas.[68] Sin emabrgo la evidencia no es conclusiva.[69] [1] Sin conocer su su dieta exacta, pero debido a su tamaño y abundancia, los iguanodontes se ven como el herbívoro dominante a grande medio para su ecosistema. [1] En Inglaterra, este incluía al depredador pequeño Aristosuchus, los grandes depredadores Eotyrannus, Baryonyx y Neovenator, pequeños herbivoros como Hypsilophodon y Valdosaurus, el también iguanodontido Mantellisaurus, el dinosaurio acorazado Polacanthus, y saurópodos como Pelorosaurus.[70]
[editar] Postura y movimiento
Reconstrucción del Iguanodon de Samuel Griswold Goodrich para Illustrated Natural History of the Animal Kingdom.[71]Los primeros fósiles eran muy fragmentarios, lo que llevo a muchas confusiones sobre la postura y naturaleza del los iguanodontes. Según lo discutido, el iguanodonte fue retratado inicialmente como bestia cuadrúpeda con un cuerno en la nariz, similar a un rinoceronte. No obstante como más huesos fueron descubiertos, Mantell observó que los miembros delanteros eran mucho más cortos que los traceros. Su rival Owen opinaba que era una criatura achaparrada con cuatro las piernas como pilares. El trabajo de supervisar la primera reconstrucción de tamaño natural de dinosaurios fue ofrecido inicialmente a Mantell, quien lo rechazo debido a su pobre salud, y a la visión de Owen formó posteriormente la base sobre la cual las esculturas tomaron forma. Su naturaleza bípeda fue revelada con el descubrimiento de los esqueletos de Bernissart. Sin embargo, fue representada en una postura vertical, arrastrando la cola a lo largo del suelo, actuando como la tercera pierna de un trípode.
Pintura del siglo XIX mostrando a Iguanodon en una pose trípode.Durante su revisión de Iguanodon, David Norman demostró que esa postura era imposible debido a la presencia de los tendones osificados a lo largo de cola.[32] Para adquirir la postura en trípode, la cola debía literalmente romperse.[5] Al poner el animal en una postura horizontal hace que muchos aspectos de los brazos y cintura pectoral sean más comprensibles. Por ejemplo, la mano es relativamente inmóvil, con los tres dedos centrales agrupados juntos, con las falanges como una pezuña y capaz de la hiperextención. Esto habría permitido que soportara el peso. La muñeca es también relativamente inmóviles, y los brazos y los huesos de hombro robustos. Estas características todas sugieren que el animal habria pasado el tiempo sobre sus cuatros miembros.[32]
Ilustración de Iguanodon como cuadrúpedo.Sin embargo los iguanodontes alcanzaban la posición cuadrúpeda en la adultez, los juveniles I. bernissartensis tenían brazos más cortos que los adultos (60% de las patas contra 70% de los adultos).[1] Al caminar como cuadrúpedo, el animal habría llevados las manos una delate de la otras, como se muestra por las vias de icnitas de iguanodóntidos y la anatomía de los brazos y manos.[72] [73] Los pies de tres dedos de los iguanodontos eran relativamente largos, y al caminar, la mano y el pie habrían sido utilizados en la manera de a digitígrada (en la punta de los dedos) .[1] LA maxima velocidad de los iguanodontes se estima en 6.7 metro por segundo, unos 24 kilómetro por hora,[74] la cuál habría sido como bípedo; no habría podido galopar como cuadrúpedo.[1]
Las huellas de pisadas de tres dedos con la punta del pie grandes se conocen en rocas del cretácico Inferior de Inglaterra, particularmente el Lecho Wealden en la isla del Wight, y éstas [[icnita] s eran originalmente difíciles de interpretar. Algunos autores asociaron a ellas con los dinosaurios a principios de 1846, E. Tagert fue más lejos en cuanto llegó a asignarlos a icnotaxón que nombró Iguanodon,[75] y Samuel Beckles noto en 1854 que se pareciás a rastos de aves, pero que pudieron haber venido de dinosaurios.[76] La identidad de los que hizieron las huellas fue aclarada con el descubrimiento en 1857 de la pierna trasera de un joven iguanodonte , con los pies que pisaban en la punta de los tres dedos, demostrando que tales dinosaurios habrían podido hacer las pistas.[77] [78] Faltando pruebas directas esta pisadas fueron atribuidasa los iguanodontes.[5] Una via de huellas de Inglaterra qué puede ser un iguanodonte lo muestra moviendoce sobre sus cuatros patas, pero las impresiones del pie son pobres, haciendo una conexión directa difícil.[32] Huellas asiganadas al icnotaxón Iguanodon se saben de localizaciones incluyendo los lugares en Europa en donde el fósil del cuerpo Iguanodon se conoce, como Spitsbergen, Svalbard, Noruega.[79] [80]
[editar] Púa del pulgar
La púa original encontrada en Maidstone en 1840.La púa del pulgar es una de la características más conocidas del los iguanodontes. Sin embargo fue colocada , en un principio, en la nariz por Mantell. Cuando se hallaron los ejemplares completos en Bernissart Dollo se dio cuenta del error y coloco en el lugar correcto en la mano, como un pulgar modificado. Ésta no sería la vez última que la garra modificada sdel pulgar de un dinosaurio sería mal interpretada; Noasaurus , Baryonyx , y Megaraptor son ejemplos desde los años de 1980 donde una garra agrandada del pulgar puesta en el pie, como en los dromeosáuridos
Este pulgar es tradicionalmente explicado como un puñal para los combates cercanos con los predadores[5] [1] aunque pudo usarlo también para romper semillass y frutas,[1] o contra otros iguanodontes.[3] Un autor propone que la garra estaba unida a una glandula venenosa,[81] pero esto no es aceptado ya que la púa no es hueca,[3] ni existe ningún tipo de estría que pudiera conducir el veneno.[82]
[editar] Comportamiento social
Aunque estén interpretados a veces como resultado de una sola catástrofe, los hallazgos de Bernissart ahora se interpreten como acontecimientos del múltiplo eventos. Según esta interpretación, por lo menos en tres ocasiones de alta mortalidad se registran, y aunque individuos numerosos habrían muerto en geológicamente un periodo breve período (10-100 años),[20] esto no es necesariamente prueba de los iguanodontes vivieran en manadas.[1] Una elemento en contra la reunión es que los restos juveniles son muy infrecuentes en este sitio, a diferencia de casos modernos con mortalidad de la manada. Eran más probablemente las víctimas periódicas de las inundaciones repentinas cuyos cuerpos muertos se acumularon en un lago o un pantano.[20] El hallazgo de Nehden, sin embargo, con su mayor abanico de edades individuales, más incluso la mezcla de Dollodon o Mantellisaurus con I, bernissartensis , y la naturaleza geográfica confinada, puede registrar la mortalidad de una manada los animales que emigran a través de los ríos.[20]
A diferencia de otros animales de manada no hay evidencia que apoyen un marcado dimorfismo sexual, como en hadrosáuridos y ceratopsios. En un tiempo, se había sugerido que I. "mantelli", o I. atherfieldensis (Dollodon y Mantellisaurus, respectivamente) de Bernissart representaban al género femenino, y los más grandes y robustos I. bernissartensis los machos.[83] Sin embargo esto no es considerado hoy en día.[32] [5] [41]
[editar] En la cultura popular
Estatua de Iguanodon por Benjamin Waterhouse Hawkins, todavia visible en The Crystal Palace, y un modelo moderno de Iguanodon del Museo de Historia Natural de la Universidad de Oxford.Desde su descripción en 1825, Iguanodon fue conocido en por la gente de todo el mundo. Dos estatuas de tamaño natural de Iguanodonse construyeron para el Palacio de Cristal en Londres en 1852 que contribuyeron en gran medida a la popularidad del género.[84] Con sus púas como cuernos en la nariz, sobre cuatro patas como elefante, sin embargo fueron la primera reconstrucción en tamaño natural de un dinosaurio
Estatua de Iguanodon en la entrada al acuario del Zoológico de Berlin.Varias películas han representado al Iguanodon. En la película de Disney Dinosaurio, un iguanodonte llamado Aladar sirvió de protagonista junto a otros tres iguanodontes y muchos otros personajes. Un paseo relatado del Animal Kingdom de Walt Disney World en Florida, Estados Unidos se trata de volver a la vida un iguanodonte. Es, también uno de los tres dinosaurios que inspiraron a Godzilla, los otros dos son Tyrannosaurus y Stegosaurus.[85] Los iguanodontes también aparecen en las series de películas de En Busca del Valle Encantado y en algunos episodios de la misma serie para televisión.
Aparte de sus apariciones en el mundo del cine, Iguanodon también se ha ofrecido en televisión la miniserie documental Caminando con dinosaurios (1999) producido por BBC, y jugado un papel estelar en el libro de sir Arthur Conan Doyle El mundo perdido. También apareció en Red Raptor (1995) de Robert Bakker, como una presa de un Utahraptor. Un asteroide del Cinturón de asteroides principal, 9941 Iguanodon en honor al género.[86] [87]
Ya que es uno de los primeros dinosaurios descritos y uno de los dinosaurios más conocidos, Iguanodon ha sido bien situado como barómetro del cambio opiniones públicas y científicas sobre dinosaurios. Sus reconstrucciones han pasado a través de tres etapas: el reptil-elefante cuadrúpedo y con un cuerno sobre el hocico de la era victoriana; un bípedo pero todavía una animal lento y torpe, que usa su cola para apoyarse y, finalmente, su representación más ágil y dinámica, en la que se desplaza tanto a dos como a cuatro patas. La segunda representación dominó el siglo XX, pero fue cambiando a partir de los años 70.[88]
Maiasauria medía entre 6 y 9 metros de longitud con un peso de casi 2 toneladas. La disposición de los huesos indica que su manera de caminar usual era bípeda, y su gran cola aplanada por los lados ayudaba a mantener el equilibrio. Si bien esta posición era útil para conseguir hojas de los árboles, seguramente se colocaban a cuatro patas para comer matorrales bajos.
[editar] Alimentación
Se alimentaba de plantas, hojas, frutas y semillas. Su hocico, con forma de pico de pato típica de los hadrosáuridos poseía varias hileras de dientes en la parte posterior de la boca que trituraban el alimento una vez se cortaba con el pico. En la parte superior del cráneo tenían una protuberancia ósea, sobre las dos cuencas oculares. Un adulto tenía que comer 90 kg de vegetales para poder tener buena salud.
[editar] Historia
Maiasaura fue descubierto por el paleontólogo de dinosaurios Jack Horner (el consejero paleontológico de las películas de Parque Jurásico) y Robert Makela en la Formación Dos Medicinas de Montana, en el año 1979. Llamó al dinosaurio Maiasaura después de una serie de nidos con restos de cáscaras de huevo, esqueletos de pequeños Maiasaura y fósiles de hojas, frutas y semillas.[1] Los nidos eran agujeros en el barro, de considerable tamaño y forma circular. Esta fue la primera prueba de dinosaurios gigantescos que cuidaban a sus crías de todo peligro hasta que sean jóvenes y puedan cuidarse solo haciendo sus propios nidos. Antes de hallar dichos nidos, estaba aceptada la idea de que no se proporcionaba ningún tipo de atención a los huevos.
Nido de Maiasaura.Cuando se continuó con las excavaciones, apenas un año después aparecieron una ingente cantidad de nidos agrupados, por lo que el lugar fue bautizado como la Montaña de los Huevos. Más de 200 especímenes, en todas las etapas de edad, han sido encontrados.
[editar] Clasificación
La relación filogenética más cercana del género Maiasaura parece corresponder con Brachylophosaurus. Para reflejar esto, ambos géneros aparecen dentro de la misma tribu. De esta manera quedan diferenciados del resto de componentes de esa subfamilia: Aralosaurus, Hadrosaurus y Gryposaurus.
[editar] Paleobiología
Debido a la gran cantidad de nidos y esqueletos encontrados en Norteamérica, los expertos creen que los maiasaura vivían en enormes manadas nómadas, que anualmente regresaban al mismo lugar de reproducción. Es posible que los nidos fueran reutilizados de un año a otro. Todas las crías que llegaran a la edad adulta, se convertirían en nuevos integrantes de la gran manada. El maiasauria fue contemporáneo al orodromeo, trodonte, el ceratópsido centrosaurio, el macizo euoplocéfalo y parientes más tempranos del tiranosaurio, Daspletosaurio y albertosaurio.
Esqueleto de un maiasaura juvenil en el Museo de Montana.Estos animales no estaban dotados con poderosas armas para repelir a los depredadores, por lo que probablemente utilizaran la huida o el camuflaje como estrategia. Es posible que poseyeran los sentidos del oído y la vista muy desarrollados para descubrir a los intrusos. Se cree que quizá pudieran esconderse sumergiéndose en lagos o ríos.
[editar] Nidos
Los maiasaurias vivían en manadas de hasta 10.000 individuos; criaban a sus jóvenes en colonias de anidamiento. Los nidos estaban muy cerca uno del otro, como en las aves modernas actuales, con un espacio entre los nidos de alrededor de 7 metros, menos que el largo de un adulto.[2] Maiasaura excavaba en el suelo un hueco de 2 m de diámetro con forma de cráter. Estos nidos estaban hechos de tierra y contenían cada uno entr 30 a 40 huevos en un patron circular o espiralado. Los huevos son de aproximadamente el tamaño de los huevos de un avestruz.[3]
Restauración de un Maiasaura con su cría.Los huevos fueron incubados por el calor resultante de la vegetación en descomposición puesta en el nido por los padres, en vez de que un padre los empollara sentandoce en el nido. Los fósiles de maiasauras bebé demuestran que sus piernas no eran completamente desarrolladas y eran incapaces de caminar. Los fósiles también demuestran que sus dientes fueron usados, así que significa que los adultos trajeron el alimento al nido.[3]
Las crías crecieron de un tamaño 16 a 58 centímetros de largo en el plazo de su primer año. A este punto, o quizás después de otro año, el animal abandonaba el nido. Esta alta tasa de crecimiento puede ser evidencia de que poseían sangre caliente. Las crías tenían diversas proporciones faciales de los adultos, con ojos más grandes y un hocico más corto.[3] Estas características se asocian a las crías y son comunes entre los animales que son dependientes de sus padres para su supervivencia durante los primeros tiempos de vida.
[editar] Instinto maternal
Reconstrucción de un maiasaura atendiendo a sus crías.Este ornitópodo cuando se apareaba, un tiempo después hacia un nido en el que ponía entre 18 y 30 huevos y esperaba hasta que nacieran. Durante la incubación, los bebés medían 50 cm de longitud y tenían huesos ligeros, extremidades largas y débiles, columna vertebral arqueada y cuello en forma de U semejante al de los adultos. Los individuos pequeños permanecían siempre en el nido porque sus patas débiles les impedían correr o caminar. Otro trabajo que tenía que encargarse la madre maiasauria era llevarles alimento al nido a los crías, las cuales no podían recolectar alimento. Sin embargo, sus dientes estaban desgastados, lo que indica un cuidado parental prolongado. Probablemente los padres masicaban las plantas duras para facilitarles el alimento a sus crías. También se ha especulado con la posibilidad que los adultos hicieran turnos de vigilancia.
[editar] Crecimiento
Cría de Maiasaura.La cría permanecía en el nido por 2 años. Los bebés crecían muy rápido, duplicaban su longitud en tan solo cuatro semanas. Las patas se volvían más fuertes y las caderas formaban una conexión con la columna vertebral. Es probable que el maiasauria joven se aventurara a alejarse del nido, en busca de alimento, bajo vigilancia de uno de los padres. Por medio de fósiles se ha descubierto que cuando alcanzaba aproximadamente 1,5 m de largo ya estaba listo para abandonar el nido y unirse a la manada. Crecían muy rápido, de hecho el crecer rápido es una de las teorías por las cuales los dinosaurios son de sangre caliente porque han necesitado alimentarse mucho para crecer de ese modo, lo que hacen normalmente los animales de sangre caliente. Ya cuando el maiasauria tenía 2 años de edad, alcanzaba longitudes de 3 m. Pero ya llegaba a la edad adulta cuando tenía entre 4 y 6 años. Las extremidades eran cortas y robustas y las patas más largas que los brazos. Al parecer la maiasauria hembra regresaba a anidar cada año al mismo lugar, hasta con 10000 animales más.[3] Cuando los jóvenes eran lo suficientemente fuertes debían migrar a otros bosques para asegurar el alimento de la manada, todas estas características se las ha afirmado por medio del hallazgo de fósiles, de esa forma se ha desifrado todo sobre su crecimiento y actitud.
De acuerdo a algunos científicos, Ankylosaurus alcanzó los 9 metros de longitud,[1] siendo uno de los anquilosaurianos más grandes que se conoce. Sin embargo, una reconstrucción sugiere un tamaño considerablemente menor, con 6,25 metros de largo, 1,5 de amplitud y 1,7 de altura a la cadera. La estructura de su cuerpo era baja y muy ancha.
Perfil del cráneo de un anquilosaurio procedente de la Formación Edmonton, Alberta.Ankylosaurus debió pesar alrededor de las 6 toneladas.[2] Fue un dinosaurio cuadrúpedo, con los miembros traseros más largos que los delanteros. Aunque no se han hallado sus pies, comparaciones con otros anquilosáuridos indican que Ankylosaurus probablemente poseía cinco dedos en cada uno. El cráneo era estrecho y triangular, más ancho que largo. El cráneo más grande que se conoce alcanzó a medir 64,5 cm de largo y 74,5 cm de amplitud. Al igual que otros anquilosaurianos, Ankylosaurus era herbívoro, y su boca se dotaba de pequeños dientes con forma de hoja que eran aptos para cortar la vegetación, los más pequeños en comparación al cuerpo de cualquier otro anquilosáurido.[3] El Ankylosaurus no compartía las series de dientes moledores de los ceratópsidos y hadrosáuridos contemporáneos, demostrando una masticación muy escasa. Tanto huesos del cráneo como en otras regiones del cuerpo, estaban fusionados para aumentar su fuerza y rigidez.[4]
[editar] Historia
El anquilosaurio fue nombrado por el paleontólogo estadounidense Barnum Brown, en 1908. El nombre genérico se deriva de las palabras griegas αγκυλος/ankulos ("curvado") y σαυρος/sauros ("lagarto"). Brown pretendió referir este nombre al término médico anquilosis, debido a su rigidez causada por la fusión de muchos huesos del cráneo y del cuerpo, por lo que el nombre es comúnmente trasladado a "lagarto rígido". La especie tipo es A. magniventris, nombre que proviene del latín magnus ("grande") y venter ("vientre"), refiriéndose a la gran anchura del animal.[5]
Restauración de Ankylosaurus sin la porra de la cola de Barnum Brown, 1908.Un equipo dirigido por Brown descubrió el espécimen tipo de A. magniventris (AMNH 5895) en la Formación Hell Creek de Montana, en 1906. Los restos consistían en un cráneo parcial, vértebras, costillas, y parte de la cintura escapular y la armadura. Seis años antes, Brown excavó el esqueleto de un gran dinosaurio terópodo (AMNH 5866) en la Formación Lance de Wyoming. El material fue nombrado Dynamosaurus imperiosus en 1905, aunque es actualmente considerado sinónimo de Tyrannosaurus rex. A este espécimen se le asignaron más de 75 osteodermos de varios tamaños, sin embargo resultan muy similares a los de A. magniventris y probablemente pertenezcan a esta especie. En 1910, durante una expedición en Alberta, Barnum Brown halló su tercer espécimen de A. magniventris (AMNH 5214), de la Formación Scollard, el cual incluía un cráneo completo y el primer mazo caudal descubierto, junto con costillas, miembros y la armadura. Los tres especímenes mencionados permanecen alojados en el Museo Americano de Historia Natural en Nueva York. El cráneo más grande conocido fue recolectado en Alberta por Charles M. Sternberg, en 1947, y ahora se mantiene guardado en el Museo Canadiense de Naturaleza. A lo largo de la historia, se han hallado numerosos huesos aislados, tales como dientes y placas óseas.[4]
[editar] Clasificación
El anquilosaurio fue nombrado como el género tipo de la familia Ankylosauridae.[5] Los anquilosáuridos son miembros del taxón mayor Ankylosauria, el cual también contiene a los nodosáuridos. La filogenia del anquilosaurio permanece discutida, con varios nuevos análisis que se contradicen entre sí, por lo que la ubicación exacta del anquilosaurio dentro de Ankylosauridae es inconclusa. Se suele considerar que el anquilosaurio y el euoplocéfalo son taxones hermanos.[1] Sin embargo, otros estudios han localizado a estos géneros en diferentes posiciones.[6] [7] Descubrimientos o investigaciones posteriores podrían aclarar este debate.
[editar] Paleobiología
[editar] Armadura
Vista dorsal del cuerpo.La característica más obvia Ankylosaurus es su armadura, la cual consiste en grandes protuberancias y placas óseas (osteodermos), incrustadas en la piel. En la piel de cocodrilos, armadillos y algunos lagartos, también se pueden identificar osteodermos. El hueso debió estar recubierto por una dura capa córnea de queratina. Estos osteodermos se diversificaron en varios tamaños, desde extensas placas semiplanas, hasta pequeños nódulos redondos. Las placas debieron de encontrarse alineadas en filas horizontales paralelas al eje central que recorrían el cuello, espalda, y caderas del animal, repitiendoce a los lados, con los pequeños nódulos protegiendo la piel entre las grandes placas. Las placas más pequeñas pudieron haberse distribuido en los miembros y la cola. Al igual que su pariente el Euoplocephalus, las placas del Ankylosaurus tenían una textura lisa, a diferencia de las altas espinas vistas en la armadura del nodosáurido contemporáneo Edmontonia. Probablemente, una fila de planas puás triangulares sobresalieron lateralmente de cada lado de la cola. El tope del cráneo estaba protegido por gruesas escamas redondas, mientras en sus esquinas posteriores, cuatro grandes cuernos piramidales se proyectaban hacia fuera.[4]
[editar] Mazo caudal
Mazo de la cola.El famoso mazo caudal del anquilosaurio también se compuso de grandes osteodermos, que se hallaban fusionados hasta las últimas vértebras caudales. El mazo era pesado y se sostenía de las últimas siete vértebras de la cola, las cuales se entrelazaban para formar una rígida vara en la base del mazo. Se han preservado gruesos tendones, los cuales se ataron a estas vértebras. Estos tendones permanecían parcialmente osificados y no eran muy elásticos, proporcionando mucha fuerza al final de la cola cuando esta se balanceaba. Al parecer el mazo funcionó como un arma defensiva, ya que pudo ser capaz de producir suficiente de un impacto devastador para llegar a romper los huesos de un atacante.[4] Se propuso también que pudo servir como un señuelo para la cabeza, sin embargo esta hipótesis ha sido muy desacreditada.[8]
[editar] Paleoecología
Ankylosaurus magniventris existió entre hace 68 y 65,5 millones de años atrás, a finales del Maastrichtiano en el Cretácico superior, siendo uno de los dinosaurios que sobrevivió hasta la extinción masiva del Cretácico-Terciario. El espécimen tipo proviene de la Formación Hell Creek en Montana, mientras otros especímenes han sido hallados en la Formación Lance de Wyoming y en la Formación Scollard de Alberta, todas de las cuales datan de finales del Cretácico.[1]
Comparación de tamaños entre un Ankylosaurus y un humano.Dichas formaciones representan diferentes secciones de la costa occidental del mar interior que dividía a Norteamérica durante el Cretácico. Se trataba de una extensa llanura costera, que recorría hacia al este desde el mar interior hasta las actuales Montañas Rocosas. Estas formaciones depositan grandes cantidades de areniscas y esquistos arcillosos, los cuales se han atribuido a ambientes de llanuras aluviales.[9] [10] [11] La Formación Hell Creek es la mejor estudiada en cuanto a los depósitos de su antiguo ambiente. En ese entonces, está región era subtropical, con un clima húmedo y cálido. La flora se diversificó rápidamente, en especial las angiospermas, que predominaron sobre las coníferas, helechos y cicadas. La abundancia de hojas fosilizadas en numerosos sitios indican que el área estaba comúnmente cubierta de pequeños árboles.[12]
A diferencia del edmontosaurio y del tricerátopo —que constituían gran parte de la fauna herbívora—, los fósiles del anquilosaurio resultan ser muy escasos en estos sedimentos. Otro anquilosauriano, edmontonia, es también hallado en las mismas formaciones. Sin embargo, el anquilosaurio y el edmontonia parecen haberse separado tanto geográficamente como ecológicamente. El anquilosaurio poseía un hocico ancho, el cual posiblemente usó para un pastoreo no-selectivo, por lo que hubiera preferido habitar en mesetas, lejos de la costa, mientras el edmontonia tenía un hocico más estrecho, sugiriendo una dieta más selectiva, y por tanto habiendo preferido las llanuras, más cerca de la costa.[4]
[editar] En la cultura popular
Desde su descripción en 1908, el anquilosaurio ha sido considerado como el dinosaurio acorazado más representativo y destacado de su grupo. Debido a su peculiar apariencia, el anquilosaurio ha llegado a ser de gran atracción en la cultura popular durante muchos años. Un modelo a tamaño real del anquilosaurio, presentado en la Feria Mundial de 1964 en Nueva York, contribuyó mucho a su popularidad.[4]
Varias películas y series han mostrado anquilosaurios. En la serie animada En Busca del Valle Encantado, hubo breves cameos de un anquilosaurio, mientras en la película Parque Jurásico III y sus videojuegos, han aparecido pastoreando junto con otros dinosaurios. En la serie Dinosaurios Extremos, hizo su aparición un anquilosaurio antropomórfo llamado "HardRock". El anquilosaurio ha sido también notable en varios documentales, tales como Paseando con Dinosaurios y La Verdad Sobre Los Dinosaurios Asesinos.
Además es notable sus apariciones en videojuegos como Zoo Tycoon, Zoo Tycoon 2 y varios juegos de Parque Jurásico. Aunque algunos de estos juegos "retratan" al anquilosaurio como un blanco difícil, otros lo caracterizan como un herbívoro débil y lento.
Corythosaurus era un herbívoro que presentaba un comportamiento gregario y se alimentaban de plantas bajas y frutos de los bosques. Medían 10 metros de largo y pesaban cuatro toneladas. Su pico no tenía dientes, pero sí la mandíbula, en la que habia cientos de pequeños y eran reemplazados constantemente a medida que caían. Estos eran usados para moler la vegetación de la que se alimentaba. Más de 20 cráneos se han encontrado de este dinosaurio. Como con otros lambeosáuridos, este animal tenia una cresta ósea alta, elaborada encima de su cráneo, que contuvo los canales nasales alargados.[2] Los canales nasales se extendieron en la cresta, primero en los bolsillos separados en los lados, luego en un solo compartimiento central y hacia adelante en el sistema respiratorio.[2] Cualquier vocalización viajaría a través de estos compartimientos elaborados, siendo amplificada.[2] Los científicos especulan que Corythosaurus podría hacer un sonido "como gritos graves como el un viento o un instrumento de latón".[2] Los sonidos serían útiles para comunicar sobre distancias largas, y podrían servir alertar el otro Corythosaurus a la presencia de alimento o a una amenaza potencial de un depredador.[2] Cuando manadas de diferentes géneros se mezclaban, su cresta, que posiblemente cambiaba de color y debe haber servido para atraer hembras y su agudo sentido olfativo debe haberle ayudado a retornar con los suyos.
Alguna vez se pensó que este dinosaurio vivió sobre todo en el agua, debido al aspecto de manos y de pies reticulados. Sin embargo, fue descubierto más adelante que el supuesto "redes" era de hecho el acolchado desinflado, como se ha encontrado en muchos mamíferos modernos.
[editar] Historia
Esqueleto completo de Corythosaurus.El primer espécimen fue descubierto en 1912 por Barnum Brown en Red Deer River, en la provincia canadiense de Alberta. Como algunos restos de coritosaurio fueron encontrados cerca de varios otros fósiles de Gryposaurus, Prosaurolophus, Lambeosaurus y Parasaurolophus, sabemos que a menudo los coritosaurios deben haberse encontrado con otros hadrosáuridos.
El mejor preservado y más completo esqueleto, hallazgo notable porque mucho del criatura se fosilizó junto con la la piel que también había sobrevivido. En 1916, el buque canadiense Mount Temple llevaba dos especímenes y otros fósiles del hoy Parque Provincial del Dinosaurio de Alberta, Canadá a Gran Bretaña. Fue hundida por la nave de guerra alemán SMS Moewe, enviando su carga de 75 millones de años al fondo del Atlántico Norte, donde se encuentra hoy en día.[3]
Se han llegado a describir siete especies, C. casaurius, C. bicristatus, C. brevicristatus, C. excavatus, C. frontalis y C. intermedius. En 1975 Peter Dodson estudió las diferencias entre los cráneos y las crestas de diversas especies de dinosaurios del lambeosaurinos. Encontró que las diferencias de tamaño y forma se pudieron haber relacionado realmente con el sexo y la edad del animal. Hoy solo se reconoce una especie, C. casuarius.[4]
[editar] Cultura popular
Corythosaurus es uno de los dinosaurios en el segmento La consagración de la primavera de la película Fantasia. Más recientemente una manada de Corythosaurus y Parasaurolophus aparecen en el film Jurassic Park III. Corythosaurus también es parte del juego de Vivendi Universal Jurassic Park: Operation Genesis.
Tuojiangosaurus (lat. "reptil de Tuojiang") es un género representado por una única especie de dinosaurio tireóforo estegosáurido, que vivió a finales del período Jurásico, hace aproximadamente 157 y 152 millones de años, en el Oxfordiano Kimeridgiano), en lo que hoy es Asia. Recobrado de la Formación Shangshaximiao en la provincia de Sichuan China.Tuojiangosaurus era bastante semejante al Stegosaurus en su aspecto y en sus hábitos alimenticios. Medía unos 7 metros de largo, 2 de alto y pesaba alrededor de 4 toneladas. Tenía quince pares de placas puntiagudas y estrechas a lo largo del dorso, desde el cuello hasta el extremo superior de la cola. Las placas más largas crecían sobre sus caderas. En el extremo de su corta cola tenía dos pares de afiladas y largas espinas; una característica también del Stegosaurus.
Extremo de la cola de un Tuojiangosaurus.
Representación de un ataque.
Esqueleto de Tuojiangosaurus.
la especie tipo y única conocida, T. multispinus, fue nombrado en 1977 (exactamente 100 años después de Stegosaurus a partir de dos ejemplares, uno con más de la mitad del esqueleto completo. Como Kentrosaurus, Tuojiangosaurus tenía dos filas de placas a lo largo de la espalda, que llegaron a ser más alta sobre la región de la cadera. También tenía dos púas a cada lado del extremo de la cola, incertadas aproximadamente a 45 grados. En los estegosáuridos, este arreglo del punto se lo ha conocido inforlamente como "thagomizer". También tenía la cabeza estrecha típica, el cuerpo arobusto, y los dientes bajos como otros estegosáuridos. Porque carecía de las altas espinas dorsales para la incercion de músculos encontrado en las vértebras del Stegosaurus, probablemente no podía alzarse en sus piernas traseras. Esto sugiere que hubiera comido la vegetación baja, cercana del suelo.[1]
Un esqueleto montado de Tuojiangosaurus multispinus se exhibe en el Museo municipal de Chongqing. Además, un modelo se ha montado en el Museo de Historia Natural de Londres. Tuojiangosaurus tambiene aparece en el especial de National Geographic Channel Dinosaurios Bizarros.
Los apatosaurios o brontosaurios eran algunos de los animales más grandes que han existido en la tierra, tenían cerca de 4,5 metros de alto hasta las caderas, una longitud total de 26 metros y una masa de hasta 24 toneladas, peso similar a 4 elefantes.[1]
La vértebra cervical y los huesos en las piernas eran más grandes y más pesados que la del diplodoco, solo que estos tenían el cuello y la cola largos. El primer cráneo fue identificado en 1975, un siglo después de que se le diera un nombre. Los apatosaurios tenía garras en sus patas, pero solamente el pulgar. Los científicos tienen la teoría de que normalmente mantenían levantada la cola un par de metros sobre la tierra. Esto evitaría que el dinosaurio la pisara, o que pusiera su cola al alcance de depredadores.
Reconstrucción de la cabeza de un apatosaurio.Se creyó que los apatosaurios eran demasiado grandes para apoyar su propio peso en tierra seca, así que se presumió que el saurópodo debió haber vivido sumergido parcialmente en el agua, o quizás en un pantano. Sin embargo, los resultados en investigaciones recientes no apoyan esto. De hecho, al igual que su primo el diplodoco, el brontosaurio era un animal de pastura con un cuello muy largo, y una cola larga que le sirvió de contrapeso. Las huellas fosilizadas indican que vivió en manadas. Para ayudar en el proceso del alimento, el Apatosaurio pudo haber tragado piedras (gastrolitos) de la misma manera que lo hacen muchos pájaros ya que sus quijadas por sí solas no eran suficientes para masticar las resistentes fibras de las plantas.
Apatosaurio.Los apatosaurio se desarrollaron en multitudes al pie de los ríos cerca de los árboles, comiendo de las hojas superiores. Los científicos creen que algunos saurópodos no podrían levantar su cuello a un ángulo de 90 grados, pues la sangre no alcanzaría a llegar a sus cabezas más de un par de minutos. Además, los estudios de la estructura de las vértebras del cuello han revelado que el cuello no era tan flexible como se pensó previamente. Nadie sabe como fue que los apatosaurios comieron suficiente alimento para satisfacer su enorme cuerpo. Probablemente comieron constantemente, deteniéndose brevemente para refrescarse, beber o para quitar parásitos. Debieron dormir mientras estaban parados. Si un depredador les atacaba, podrían defenderse azotando su cola de lado a lado, o pisando sobre su depredador. Debido a la lenta velocidad en la que se desplazaban los apatosaurios, estos vivieron en manadas, y podían "llamarse" uno a otro, en caso de requerir ayuda.
[editar] Sistemática
Apatosaurus es un miembro de la familia Diplodocidae, que también incluye a dinosaurios como Diplodocus y Barosaurus, aunque no sea tan estrechamente vinculado a estos géneros como lo están el uno al otro, y por lo tanto Apatosauruses usualmente ubicado en su propia subfamilia, Apatosaurinae, junto con sus cercanos relativos Supersaurus y Eobrontosaurus.[2] [3] [4]
En 1877, Othniel Charles Marsh publica el nombre de la especie tipo Apatosaurus ajax. Continúa en 1879 con la descripción de otro espécimen más completo, que él pensó que representaba un nuevo género y lo llamó Brontosaurus excelsus. En 1903, Elmer Riggs argumenta que Brontosaurus excelsus era tan similar a Apatosaurus ajax que pertenece al mismo género, y junto con Riggs lo reclasificó como Apatosaurus excelsus. De acuerdo con las reglas de ICZN el nombre Apatosaurus, fue el primero en publicarse y dejando a Brontosaurus como sinónimo y descartándolo para su uso oficial.
Diplodocidae
Apatosaurinae
Suuwassea
Supersaurus
Apatosaurus
Diplodocinae
Barosaurus
Diplodocus
Cladograma de Diplodocidae por Lovelace, Hartman, y Wahl, 2008.[4]
[editar] Especies
[editar] Apatosaurus ajax
Es el espécimen tipo de este género, y fue nombrado por el paleontólogo Othniel Charles Marsh en 1877 en honor a Ajax, el héroe de la mitología griega. Se conoce por el holotipo y dos esqueletos parciales incluyendo parte de un cráneo.
[editar] Apatosaurus excelsus
Originalmente Brontosaurus fue nombrado por Marsh en 1879. Se han encontrado restos de esta especie en los Estados Unidos, parte de seis esqueletos parciales, incluyendo piezas de un cráneo, en Colorado, Oklahoma, Utah y Wyoming.
antigua representación de A. (Brontosaurus) excelsus.[editar] Apatosaurus louisae
Fue nombrado por William Holland, en 1915 en honor de señora Louise Carnegie, esposa de Andrew Carnegie quién financió la investigación de campo para encontrar los esqueletos completos del dinosaurio en el oeste americano. Apatosaurus louisae se conoce a partir de un esqueleto parcial que fue encontrado en Colorado, Estados Unidos.
[editar] Apatosaurus parvus
Apatosaurus parvus originalmente Elosaurus parvus, pero reclasificado como una especie de Apatosaurus en 1994.[5] Esta sinonimia fue mantenida en 2004.[6]
[editar] Especies inválidas
Robert T. Bakker clasificó al tipo de “Apatosaurus yahnahpin” como la especie de un nuevo género en 1998: el Eobrontosaurus. Así que ahora es apropiadamente denominado como Eobrontosaurus yahnahpin. Fue nombrado por Filla, James y Redman en 1994. Un esqueleto incompleto ha sido descubierto en Wyoming. Sin embargo, se ha discutido recientemente si Eobrontosaurus pertenece a Camarasaurus.[7] En 1957 Lapparent y Zbyszewski nombraron a A. alenquerensis en Portugal, hoy conciderada Lourinhasaurus. A. minimus Mook, 1917 y A. montanus Marsh, 1877/von Huene, 1906 ambos dudosos en incluidos en Atlantosaurus. A. grandis Marsh, 1877 es hoy considerado Camarasaurus sp. A. amplus Marsh, 1881/Gilmore, 1936 es un sinónimo menor de A. excelsus.
[editar] Palaeobiología
Fémur de Apatosaurus en Cosmocaixa, Barcelona.En un principio se pensó que los apatosaurios eran demasiados masivos para soportar su propio peso sobre terreno firme, por lo que teorizo que estos sarópodos debían vivir en pantanos parcialmente sumergidos en el agua. Desde ese momentos los descubrimientos, no solo no apoyaron esta idea, es más hoy se cree que al igual que sus parientes los diplodocos, los apatosaurios vivían sobre suelo seco pastoreando, con su largo cuello y cola actuando como contrapesos. Un estudio ha descubierto que los diplodócidos tenían cuellos mucho menos flexible que lo que se pensaba, no lo llevaban erguido como un cisne, sino en línea recta paralelo al suelo, de donde se alimentaban.[8] Las icnitas encontradas sugieren que vivían en manadas.
En 2008, huellas de un apatosaurio juvenil fue reportada en la Mina Five de Morrison, Colorado. Descubierta en 2006 por Matthew Mossbrucker, estas pisadas demuestran que los apatosaurios jóvenes erán capaces de correr sobre sus patas traseras, asemejándose a un lagarto basilisco.[9]
[editar] Postura
A principios del siglo XX, los diplodócidos, como los apatosauriso, eran retratados con sus largos cuellos erguidos, alimentándose de la vegetación de las copas de los árboles. Más recientementes, los científicos han sostenido que el corazón no habría podido crear la suficiente presión arterial para abastecer de oxigenar el cerebro. Además, estudios más recientes han demostrado que la estructura de las vértebras del cuello no habría permitido que el cuello se doblara mucho hacia arriba.[10] [11]
[editar] Fisiología
Con una masa tan grande del cuerpo, combinada con un cuello largo, los fisiólogos encuentran problemas para determinar cómo estos animales pudieron respirar. Comenzando por asumir que los apatosaurios, como los cocodrilos, no tenían diafragma, el volumen del espacio muerto (la cantidad de aire que permanece en la boca, la tráquea y los conductos de aire después de cada respiración) se ha estimado aproximadamente en 184 litros para un espécimen de 30 toneladas. El cálculo de su capacidad pulmonar (la cantidad de aire movida adentro o hacia fuera durante un único movimiento de respiración) se basó en la presunción de distintos modelos de sistemas respiratorios:
904 litros si es similar a las aves .
225 litros si es similar a los mamíferos.
19 litros si es similar a los reptiles.
Sobre esta base, su sistema respiratorio no habría podido ser el propio de un reptil, pues su volumen de aire inspirado no habría podido sustituir el volumen en el espacio muerto. Asimismo, el sistema mamífero proporcionaría solamente una fracción de aire nuevo en cada respiración. Por lo tanto, debe haber tenido un sistema desconocido en el mundo moderno o uno similar al de las aves, es decir sacos aéreos múltiples conectados con los el pulmones. Además, un sistema aviar necesitaría solamente una capacidad pulmonar de cerca de 600 litros comparados a un requisito mamífero de 2.950 litros, que excederían el espacio disponible. El volumen torácico total de los apatosaurios se ha estimado en 1.700 litros teniendo en cuenta 500 litros, de un corazón de cuatro cámaras (como las aves y cocodrilos, no de tres como los reptiles) y una capacidad pulmonar de 900 litros. Eso permitiría cerca de 300 litros para oxigenar el tejido. Presumiendo que los apatosaurios tenía un sistema respiratorio aviar y un metabolismo de reptil, necesitarían consumir solamente cerca de 262 litros de agua por día.[12] No se sabe cómo los apatosaurios podían comer suficiente alimento para satisfacer las necesidades de sus enormes cuerpos, pero es probable que comieran constantemente, deteniéndose brevemente para refrescarse, beber o quitarse parásitos. Se conjetura que dormían de pie. Confiaron probablemente en su tamaño y en el número de sus manadas para disuadir a los depredadores.
[editar] Cola
Un artículo de noviembre de 1997 en la revista Discover Magazine comenta la investigación de la mecánica de la cola de los apatosaurios, realizada por Nathan Myhrvold, un informático de Microsoft. Myhrvold realizó una simulación de computadora de la cola, que en diplodócidos como los apatosaurios tenían una estructura muy larga, con un ahusamiento que se asemejaba a un látigo. Este modelado de la computadora sugirió que los saurópodos eran capaces de producir con ella un sonido similar a un estallido de alrededor de 200 decibelios, comparable al volumen del sonido de un cañón.[13]
[editar] Historia (el brontosaurio)
Apatosaurus significa lagarto engañoso, fue dado porque los huesos del galón eran similares a los de un lagarto marino prehistórico, Mosasaurus. Apatosaurus proviene del griego ἀπατέλος o ἀπατέλιος (apatao) engañoso y σαῦρος (saurus) lagarto. Los fósiles de este animal se han encontrado en Mina Nine Mile y Mina Bone Cabin en Wyoming y en los sitios en Colorado, Oklahoma y Utah, los EE.UU..
Diagrama del esqueleto realizada por Marsh en 1896.En 1877, Othniel Charles Marsh publicó notas sobre el descubrimiento del Apatosaurus, y entonces en 1879 describió otro hallazgo mucho más completo como Brontosaurus. Marsh nombro la nueva especie como Brontosaurus excelsus, que significa el " lagarto del trueno", del griego brontē/βροντη traducido como trueno y sauros/σαυρος como lagarto y del latín excelsus, "excedido en gran número", refiriendo al mayor número de vértebras sacras que en cualquier otro género del saurópodo conocido en ese entonces. Pero durante 1903 fue descubierto que el Apatosaurus era de hecho un Brontosaurus joven, y siendo que el nombre de Apatosaurus fue publicado primero, se juzgo que este tenía prioridad como nombre oficial; entonces el término Brontosaurus fue relegado a ser un sinónimo menor. El nombre Brontosaurus no fue formalmente eliminado de los expedientes de la paleontología hasta 1974.
Esqueleto de Apatosaurus.Fue el dinosaurio más grande y completo conocido en ese entonces, careciendo solamente de una cabeza, pies, y las porciones de la cola y fueron preparados para cuál era ser la primera exhibición montada de un esqueleto del saurópodo, en el Museo Peabody de Historia Natural de Yale en 1905. Los huesos que falta fueron creados usando pedazos conocidos de familiares cercanos de Brontosaurus. Los pies fueron agregados de saurópodo que se descubrieron en la misma mina, también una cola formada para asemejarse a la que Marsh tenía, además de un modelo compuesto de lo que él pensaba que sería el cráneo de esta criatura masiva. Este no era el de un delicado Diplodocus,[14] sino el compuesto "con huesos más grandes, más gruesos, más fuertes en el cráneo, mandíbulas inferiores y las coronas del diente a partir material proveniente de tres diversas locaciones",[15] primariamente del Camarasaurus, el único el otro saurópodo del cual se conocía buen material del cráneo en ese entonces. Este método de reconstruir los esqueletos incompletos basados en los restos más completos de dinosaurios relacionados continúa en montajes del museo y restauraciones hasta la actualidad.
A pesar del principio mucho de lo publicado del esqueleto montado, que cementó el nombre Brontosaurus en el público, Elmer Riggs había publicado un trabajo en 1903 en Geological Series of the Field Columbian Museum sosteniendo que Brontosaurus no era lo bastante diferente de Apatosaurus para autorizar su propio género, y creado la combinación Apatosaurus excelsus.
… debido a estos hechos los dos géneros se pueden ver como sinónimos. Como término Apatosaurus tiene prioridad sobre Brontosaurus; este último será considerado sinónimo.
A pesar de esto, por lo menos un paleontólogo Robert Bakker; sostuvo en los años 1990 que A. ajax y A. excelsus son de hecho suficientemente distintos para continuar mereciendo un género separado.[16] Esta idea no ha sido aceptada por la comunidad científica.[7] [17]
[editar] En la cultura popular
El tiempo transcurrido para reparar la clasificación errónea de Marsh causó que el nombre Brontosaurus se hiciera popular, y fuera asociado con uno de los dinosaurios más grandes; llegó a ser tan famoso que persistió de largo después de que el nombre hubiera sido abandonado oficialmente en uso científico. El brontosaurio, los brontosaurios, y los brontosaurianos de los términos de uso frecuente se usan para referirse genéricamente a los saurópodos.
Imagen de apatosaurios en un pantano, realizada a principios del siglo XX, realizada con lo que se presumía en esa época.Aun en 1989, el Servicio Postal de los Estados Unidos emitió una serie de estampillas con cuatro dinosaurio, Tyrannosaurus, Stegosaurus, "Pteradon" (error ortográfico por Pteranodon, que es un pterosaurio y no un dinosaurio) y Brontosaurus. La inclusión de estos dos últimos levantó las quejas sobre sus errores científicos.[18] A lo que el servicio de correo se defendió arguyendo: Aunque ahora sea reconocido por la comunidad científica como Apatosaurus, el conocido "Brontosaurus" fue utilizado para la estampilla porque es más familiar al público en general. Stephen Jay Gould apoyó esta posición en su ensayo "Bully for Brontosaurus", aunque él repitió la discusión original Riggs que " Brontosaurus" es un sinónimo con el "Apatosaurus". Sin embargo, él observó que la criatura ha desarrollado y continúa manteniendo una existencia independiente en la imaginación popular.[19] En el documental de la BBC Walking with Dinosaurs, en el especial "La Balada del Gran Al" (The Balad of Big Al), aparece una manada de apatosaurios, siendo nombrados de esa manera y no con el más popular brontosaurio. Desde el principio de la historia del cine, Brontosaurus se ha representado en película. Por ejemplo, la película muda de1925 El mundo perdido, se lo vio gracias a los efectos especiales de Willis O' Brien, en una batalla entre un Brontosaurus y un Allosaurus . En King Kong de 1933 se vio un Brontosaurus, al igual que en la remake de 2005, fue descrito usando el nombre científico ficticio; Brontosaurus baxteri. El brontosaurio de la película 2005 es perceptiblemente diferente de la visión moderna del Apatosaurus, con una cabeza cuadrada, una cola baja colgante, y un cuello de tipo serpiente evocador de las pinturas del período de los años 30 de la especie.
Protoceratops llego a medir 1,8 metros de largo, 60 centímetros de alto hasta la cruz y un peso 180 kilogramos. El gran número de estos aimales encontrados en grandes concentaraciones hace pensar que vivieran en manadas.[2] Protoceratops estaban emparentados con el Leptoceratops, un protoceratopsido que habitaba en Norteamérica. Algunas revisiones, como la de Peter Dodson, atribuyen la diferencia de tamaño y forma de algunos huesos craneales - concretamente el parietal y el escamoso - a la existencia de cierto dimorfismo sexual, además de la edad.[3] Su aspecto debió ser intimidatorio a pesar de su relativo pequeño tamaño, ya que su cabeza estaba protegida por una maciza placa y su boca poseía un afilado pico. La cola tenía una longitud considerable y era gruesa. La reconstrucción del esqueleto indica que este animal era cuadrúpedo. La placa, además de proteger de posibles ataques frontales, probablemente fuera un elemento más del cortejo durante el periodo de apareamiento. Los músculos de la mandíbula eran prominentes, capaces de realizar una gran fuerza. De esta manera era capaz de arrancar las plantas y hojas de las que se alimentaba. Después masticaba los alimentos con sus hileras de fuertes dientes.
Protoceratops era un relativamente pequeño dinosaurio con una enorme cabeza. Aunque fue un herbívoro, Protoceratops parece haber tenido una poderosa mordida. Estas mandíbulas tenían grandes baterías de dientes bien adaptados para masticar la vegetación resistente.[2] El cráneo terminaba en un pico frontal muy fuerte y estba perforada en cuatro pares de ventanas . El primer agujero era las narinas, que era la considerablemente más pequeño que las vistas en los géneros posteriores. Protoceratops tenía una órbita ocular grande, que midia alrededor de 50 milímetros de diámetro.[4] Detrás del ojo era una ventana levemente más pequeña, la fenestra infratemporal.
Este dinosaurio cuadrúpedo tenia una gola ósea en la paarte posterior de la cabeza. La gola en si estaba perforada por dos grandes fenestras parietales, en las mejillas sobresalian grandes huesos yugal.[5] El tamaño exacto y la forma de la gola del cuello variaron en cada individuo, algunos especímenes tenían golas cortos y compactas, mientras que otros las tenían de una longitud de casi mitad del cráneo. La gola consiste sobre todo en el parietal el y parcialmente en el escamoso. Algunos investigadores, includidos Peter Dodson[4] atribuye las diferencias de tamaños y formas de los huesos a el dimorfismo sexual y a la edad del especimen.
[editar] Historia
Protoceratops y Velociraptor en combate.En 1922 una expedición científica que buscaba a los ancestros humanos en el desierto de Gobi de Mongolia, encabezada por el explorador estadounidense Roy Chapman Andrews, encontro los primeros restos. Fue el fotógrafo, J.B. Shackelford, del grupo el que consiguió encontrarlos. Además, se encontrarón los primeros huevos de dinosaurios . Este descubrimiento demostró por primera vez que los dinosaurios eran ovíparos, anteriormente, aunque se considerara que los dinosaurios eran reptiles, no se sabía como estos animales se reproducían. En un nido se encontraron cerca de una treintena de huevos, por lo que se pensó que varias hembras de Protoceratops usaban el mismo habitáculo. Durante mucho tiempo se pensó que el Oviraptor robaba los huevos de estos animales, sin embargo, en 1993 se descubrió un embrión de Oviraptor dentro de esos supuestos huevos de Protoceratops. Aunque no se encontraron fósiles de humanos primitivos, la expedición, recolecto muchos especimenes de Protoceratops junto a los restos de los terópodos: Velociraptor, Oviraptor, y el más primitivo aun ceratopsiano Psittacosaurus.[4]
Recreación artística de un velocirráptor y un protoceratopo enzarzados en un combate mortal, basándose en los especímenes de los "Dinosaurios Luchadores".Walter Granger y W.K. Gregory describieron formalmente la especie tipo, P. andrewsi en 1923, el nombre especifico es en honor a Andrews. Los restos provenían de la Formación Minhe, Gansu, Mongolia interior] Zizhiqu, China. Los investigadores inmediatamente notaron la importancia de los descubrimientos de Protoceratops, y el género fue descrito como el "tan buscado ancestro de Triceratops".[4] Los fósiles se encuentran en excelente estado, con el anillo esclerotico, unos delicados huesos de la orbita ocular, preservados en algunos de los especimenes.[4]
En 1971, un fósil de este dinosaurio fue encontrado entrelazado en combate con un Velociraptor en la Formación Djadochta en Omnogov Mongolia. Se cree que los dos murieron simultaneamente, mientras peleaban, cuando o fueron sorprendidos por una tormenta de arena o enterrados cuando una duna se derrumbó encima de ellos.[6]
El 1975, los paleontólogos polacos Teresa Maryanska y Halszka Osmólska describieron una segunda especie de Protoceratops, del Campaniano de Mongolia, al que llamaron P. kozlowskii.[7] Sin embargo, consistían en restos incompletos de un ejemplar juvenil, y hoy en considerada sinónimo menor de Bagaceratops rozhdestvenskyi.[8]
En 2001, una segunda especie válida, P. hellenikorhinus, fue nombrada proveniente de la Formación Bayan Mandahu en Mongolia interior, China y también es datada del Campaniano. Esta es probablemente tan larga como P. andrewsi, teniendo una significativamente distinta gola, teniendo cuernos yugales más robustos. El arco del hueso sobre sus ventanas de la nariz tenía dos pequeños cuernos nasales, y no había dientes en el frente del hocico.[9]
[editar] Teoría sobre el origen del mito del grifo
La investigadora del folklore y de la historia científica, Adrienne Mayor de la Universidad de Stanford, ha sugerido que los fósiles magníficamente preservados de Protoceratops y otros dinosaurios con pico podrían ser la base para el mito del grifo. Los antiguos nómadas escitas que extraían oro en los montes Altai y Tien Shan de Asia Central, pudieron encontrar casualmente algún resto de estos animales y elucubrar a partir del pico que se trataba de enormes bestias voladoras, a los que bautizaron como grifos. Los grifos son descritos como cuadrúpedos del tamaño de un leon, con enormes garras y un pico similar al de aves rapaces, que ponían sus nidos en nidos en el suelo.[10]
Los escritores griegos comienzan a describir al grifo alrededor de 675 A.C., al mismo tiempo que tuvieron contacto con los escitas. Los grifos fueron descritos como guardianes de los depósitos del oro en las colinas áridas y las formaciones de la piedra arenisca roja de las tierras yermas. Las montañas vecinas de la región de Mongolia y China de donde se encuentran muchos fósiles de Protoceratops son ricas en oro, dando cierto crédito a la teoría que estos fósiles eran la base de los mitos del grifo.[4]
[editar] Clasificación
Protoceratops fue el primer protoceratópsido nombrado y quien le diera el nombre a la familia Protoceratopsidae, un grupo de dinosaurios herbívoros que fueron muy avanzados también para ser psitacosáuridos, pero primitivos a ser ceratópsidos. Semejantes a Ceratopsidae pero con más caractericas de grupo cursorial en las proporciones del miembro, golas generalmente más pequeños, y carencia de cuernos grandes.
En 1998, Paul Sereno definió a Protoceratopsidae como el clado nodo vástago que incluye a "todos los coronosaurios mas cercanos a Protoceratops que a Triceratops." Algunos estudios ubican a Bagaceratops, Breviceratops, Graciliceratops, Lamaceratops, Magnirostris, Platyceratops y Serendipaceratops dentro de Protoceratopsidae, pero en 2006, Makovicky y Norell publicaron una nueva filogenia en la que quitan a muchos géneros de Protoceratopsidae, además de varias otras filogenias que también existen. Bainoceratops podría ser considerado como sinónimo menor de Protoceratops.[11]
[editar] Sistemática
Coronosauria Leptoceratopsidae
Montanoceratops
void
Udanoceratops
Leptoceratops
Prenoceratops
void
Graciliceratops
void Protoceratopsidae
Bagaceratops
Protoceratops
void
Zuniceratops
Ceratopsidae
[editar] Paleobiología
[editar] Reproducción
Un cráneo embrionario de Protoceratops.En la década del 1920, Roy Chapman Andrews descubrió los primeros huevos fósiles de dinosaurios, en el Desierto de Gobi de Mongolia. Cada huevo media alrededor de 20 centímetros de diámetro y la cría recién nacida debía haber medido 30 centímetros de largo. Debido a la proximidad con los fósiles de Protoceratops, estos huevos se asignaron a este género.
Cría deProtoceratops'.se creyó que el terópodo contemporáneo Oviraptor se alimentaria de huevos de Protoceratops debido a un esqueleto de Oviraptor presente en el nido. El cráneo estaba aplastado, y se especulo que la lesión fue recibida por un enojado Protoceratops que defiendia su nido del depredador. Sin embargo, en 1993 Norrell et al. Un embrion dentro del supuesto huevo de Protoceratops. Este embrión, luego de la reexaminación más cercana, resultó pertenecer realmente a Oviraptor.[12] El hallazgo original realmente representa un comportamiento de empollamiento de Oviraptor más que un intento fallido de comer los huevos.
Igualmente se han encontrado agrupaciones de nidos de Protoceratops, por lo cual es posible que los Protoceratops vivieran en manada o con una organización de tipo familiar. Al nacer, estos animales tenían una longitud de 30 centímetros aproximadamente, y eran alimentados hasta que eran capaces de tener cierta autonomía.
aki kreo ke termina mi obra adios.